The Project Gutenberg EBook of Comparative Breeding Behavior of Ammospiza caudacuta and A. maritima, by Glen E. Woolfenden This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: Comparative Breeding Behavior of Ammospiza caudacuta and A. maritima Author: Glen E. Woolfenden Release Date: May 30, 2011 [EBook #36285] Language: English Character set encoding: UTF-8 *** START OF THIS PROJECT GUTENBERG EBOOK COMPARATIVE BREEDING *** Produced by Chris Curnow, Joseph Cooper and the Online Distributed Proofreading Team at http://www.pgdp.net
University of Kansas Publications
Museum of Natural History
Vol. 10, No. 2, pp. 45-75, 6 pls., 1 fig.
December 20, 1956
Comparative Breeding Behavior of Ammospiza caudacuta and A. maritima
BY
GLEN E. WOOLFENDEN
University of Kansas
Lawrence
1956
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Harrison B. Tordoff
Volume 10, No. 2, pp. 45-75, 6 pls., 1 fig.
Published December 20, 1956
University of Kansas
Lawrence, Kansas
PRINTED BY
FERD VOILAND. JR., STATE PRINTER
TOPEKA, KANSAS
1956
Comparative Breeding Behavior
of Ammospiza caudacuta and A. maritima
BY
GLEN E. WOOLFENDEN
CONTENTS
| PAGE | |
| Introduction | 48 |
| Materials and Methods | 48 |
| Description of the Area | 49 |
| Flora | 50 |
| Reptiles | 50 |
| Mammals | 50 |
| Predators | 50 |
| Passerine Associates | 51 |
| Winter Status and Spring Migration | 51 |
| Territory | 52 |
| Voice | 58 |
| Song | 58 |
| Calls | 60 |
| Copulation | 61 |
| Nests | 62 |
| Eggs and Incubation | 65 |
| Young | 65 |
| Growth | 65 |
| Behavior | 68 |
| Food, Feeding and Care of the Young | 71 |
| Acknowledgments | 73 |
| Summary | 73 |
| Literature Cited | 74 |
INTRODUCTION
Taxonomically the Genus Ammospiza has received the attention of ornithologists for more than a century. Nevertheless, the behavior of no species of the genus has been studied extensively. The papers of Montagna and Tomkins are the only works that mention behavior and natural history in any detail. There has been an increasing awareness of the importance of ethological data and of their usefulness in systematics. For these reasons, I made a comparative study of the breeding behavior of the Sharp-tailed Sparrow (Ammospiza caudacuta) and the Seaside Sparrow (Ammospiza maritima) in New Jersey in the spring and summer of 1955.
The Seaside Sparrow is restricted to the Gulf- and Atlantic-coasts of North America, breeding north to Massachusetts. The Sharp-tailed Sparrow breeds south to North Carolina. The overlap of the breeding ranges of the two species is therefore small. Furthermore the forms breeding in the coastal states are restricted to tidal marshes, and the geographically peripheral colonies of each species are small. Irregular nesting is the case for the northernmost colonies of the Seaside Sparrow, on Cape Cod (Griscom, 1944:317), and the same is probably true for the colonies of the Sharp-tailed Sparrow on Pea Island, North Carolina, as indicated by Montagna's failure to locate any breeding birds in July, 1941 (Montagna, 1942b: 256). The center of overlap of the ranges of the two species is in New Jersey where both forms are abundant and can best be studied comparatively.
MATERIALS AND METHODS
The adult sparrows were captured and banded, and sometimes the nestlings were banded. The standard funnel trap, baited with seeds, proved useless for capturing birds of the Genus Ammospiza, although migrant Savannah Sparrows (Passerculus sandwichensis) readily entered. A Japanese bird net, twenty-five feet long, was used successfully. Eighty-five Sharp-tailed Sparrows and forty Seaside Sparrows were banded at two localities.
All of the adult sparrows were banded with United States Fish and Wildlife Service numbered bands and colored celluloid bands. The colored bands I used were obtained from the Hinton Supply Company of New York City, which manufactures them for cage birds. The firm makes them in seven colors, sold at reasonable prices. With seven colors, the number of combinations, using only one colored band and one aluminum band per bird, is forty-two.
In addition, I dyed many adults and all nestlings. Alcoholic solutions of Victoria Blue B S concentrate and Alizarine Red S concentrate were used. The males were dyed red, the females blue; various areas of the body were colored in order further to individualize the birds. Although the dyes disappeared in less than a month, the markings were helpful on many occasions.
When an adult bird was captured I always sexed it and ordinarily weighed and measured it. The nestlings were weighed and measured daily at intervals of 24 hours. I built a corral of hardware cloth around one Sharp-tailed Sparrow nest in order to measure the young after they left the nest. The sex of any adult was ascertained by examining the cloacal area, as described by Salt (1954:61-75). Sex as determined by this method was corroborated by internal examination of the specimens collected.
A pan balance accurate to one-tenth of a gram was used for weighing. The adults were weighed in a cloth sack, the sack being weighed each time to prevent error owing to variable moisture and other factors.
Dragging the marsh with a rope was ineffectual in finding nests. The birds flushed long before the rope neared them. I found nests of the sparrows by using a blind. From a blind I would determine the approximate location of a nest by watching the movements of the adult birds. Then I would either make a direct search of the vegetation or move the blind closer to find the actual site.
Many hours were spent in blinds. I had two in operation throughout the breeding season, and it was from these that most of the data on behavior were accumulated. Observations were made by means of a 7 x 50 coated binocular and on occasions by means of a 19.6x spotting telescope.
DESCRIPTION OF THE AREA
The intensive work was carried out on the marshes west of the town of Lavallette in Ocean County, New Jersey. Further observations were made at other localities in the county, in particular at the Chadwick marshes (plate 6), one mile north of the Lavallette site, where many of the Ammospizas were banded. The breeding Ammospizas of the localities are the nominate races, A. c. caudacuta (Gmelin) and A. m. maritima (Wilson).
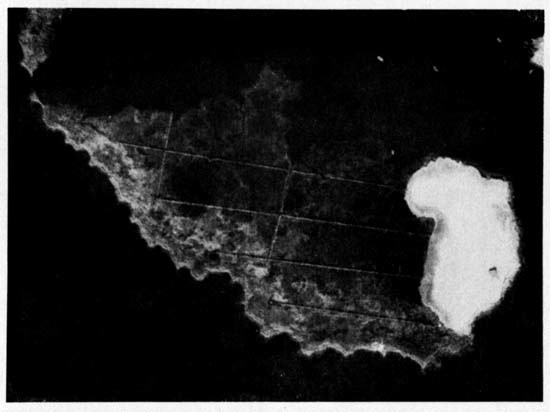
Characteristic of the sand beaches of the Atlantic coast of the United States are offshore bars which, when exposed, form long bays parallel to the coastline. These bays become surrounded by marshes that in turn are inhabited by the two species of Ammospiza. The birds prefer the marshes closest to the ocean (plate 6). I made trips to the marshes on the mainland side of upper Barnegat Bay and found only a few Sharp-tailed Sparrows and no Seaside Sparrows in residence.
The island of the Lavallette marshes that I worked on was approximately 1400 feet long and 600 feet wide. One-third of the east central end of the island was covered with sand fill, pumped there several years before the study was begun (plate 1, fig. b). The island was also ditched. The four east-west ditches are spaced 125 feet apart; the two ditches perpendicular to these are 340 feet apart and are situated in the western portion of the island. These ditches, originally dug as a means of decreasing the mosquito population, are one foot wide and almost three feet deep. The excavated earth is deposited in a row paralleling the ditch. The entire island, excluding the sand fill is not more than two feet above normal high tide. In August, 1955, abnormally high water, a result of hurricane "Connie", rose four to five feet and covered all but the tops of the bushes and a few mounds of sand. Low tides expose no mud flats for the edges of the marsh are nearly vertical banks and the water along the edges is more than one foot deep.
The average temperature for July, compiled over a 34 year period at the Asbury Park weather station is 72.6°F. The average precipitation from May through August, acquired over the same length of time, is between 3.5 and 4.5 inches per month.
In spring and summer the prevailing winds are from the south and southwest. Therefore, the south and west shores of the island are subject to greater inundations by water. The fact that the island is unprotected by neighboring islands from the open expanse of the bay on this side is also of importance in this respect. The north and east shores, on the lee side of the island, are guarded from the open bay by nearby land. The exposed southern shores, where there was open mud and sparse patches of cord-grass, were the preferred feeding areas of the Seaside Sparrows. Lack of exposed and open feeding areas may account for the absence of this species in areas that otherwise seem to fulfill the requirements of the species.
Two major drift lines were present on the island: one within a few feet of the waterline consisted mostly of dead eel grass (Zostera marina), and the other, situated close to the cattail strip, contained a variety of flotsam (pl. 2, fig. a).
FLORA
The vegetation on the island consisted chiefly of smooth cord-grass (Spartina alterniflora), black grass (Juncus gerardi), cattail (Typha sp.), and marsh-elder (Iva frutescens). Other plants identified on the area were: common reed grass (Phragmites communis) and slender grass wort (Salicornia europea). Black grass grows on the inner, dryer portions of the marsh, and cord-grass prefers the wetter portions, growing to the edge of the water. The marsh-elder bushes mostly are restricted to the mounds of earth dug from the ditches. Cattails, in general, grow in a narrow band paralleling, but back a few yards from, the shoreline. Areas of mixed black grass and cord-grass occurred.
REPTILES
Diamond-backed terrapins (Malaclemys terrapin) were the only reptiles recorded from the study island. Several were taken on land, but the majority were seen in the waters about the marsh.
On June 27 a black snake (Coluber constrictor) was seen in a bushy area bordering a marsh on the mainland side of Barnegat Bay. A few Sharp-tailed Sparrows were seen in the same locality and a singing male (G. E. W. 559) with testes 14 x 8 mm. and a female (G. E. W. 558) with a brood patch were collected.
MAMMALS
Only two species of mammals, both abundant, were present on the study island: the meadow vole (Microtus pennsylvanicus) and the muskrat (Ondatra zibethicus). The muskrats dug burrows beneath the level of the water into the banks of the island, used the ditches as routes to the interior of the marsh and built some small houses, mostly from cattail stems.
PREDATORS
Unless the above named mammals preyed on the sparrows, all of the enemies of the colony at Lavallette were avian. Both Crows (Corvus brachyrhynchos) and Fish Crows (Corvus ossifragus) visited the local marshes frequently as did a Marsh Hawk (Circus cyaneus). I watched the Marsh Hawk make many passes at what I thought were sparrows, but the only animal I ever saw caught by the hawk was a Microtus. The sparrows were alarmed when the hawk appeared, quickly and silently disappearing into the grass.
At least two nests on the Lavallette Marsh were destroyed by predators in the course of the breeding season of 1955. One nest of the Seaside Sparrow was empty when I checked it on July 3; on July 2 it had contained four young, three days old. On July 21 I found a dead Sharp-tailed Sparrow, approximately three days old, lying on a patch of matted grass. A hole was in the flank of the bird and blood was present about the bill. This nestling was not from a nest under observation.
PASSERINE ASSOCIATES
On the Lavallette marshes the only passerine associates of the two species of Ammospiza were Song Sparrows (Melospiza melodia atlantica) and Long-billed Marsh Wrens (Telmatodytes palustris palustris). Two pairs of Song Sparrows and less than six pairs of marsh wrens nested on the study area. One Song Sparrow nest was found and is plotted on the map (pl. 1, fig. b); the other pair nested somewhere along the east shore of the island. The Song Sparrows at the east end of the island obtained most of their food from the grounds of the Lavallette Yacht Club across fifty yards of water to the east. The pair that nested in the western portion of the island fed along the sand fill or along the bases of the marsh-elder. Their nest was built in the most extensive area of these bushes; it was placed approximately one foot above the ground in a small dead bush and gained support and concealment from the surrounding black grass. Three of the four eggs hatched on June 30, and the young left the nest on July 11. Both parents fed the offspring.
The marsh wrens fed and nested in the cattails. I never saw these wrens away from the cattails.
WINTER STATUS AND SPRING MIGRATION
Ocean County is ten miles south of the area treated in Cruickshank's regional work (1942). He considers both species as rare to casual winter residents. Concerning the spring migration of the Sharp-tailed Sparrow he says (p. 456) "The first widespread wave never comes before April 25, however, and most of the birds arrive in May." He mentions that late May is the height of migration and that stragglers are passing through up to the middle of June. The arrival of the Seaside Sparrow in spring is similar (p. 458): the first widespread movement is in early May, the peak is reached in the third week of the month, and stragglers have been recorded through the second week in June.
I was in the field in Ocean County almost daily all spring and found no Seaside Sparrows and only two Sharp-tailed Sparrows north of Barnegat Inlet, Ocean County, before May 5. I waded through the marshes at Chadwick, Lavallette, and Island Beach State Park on April 27 when high tides covered all of the dense vegetation and saw no sparrows of the Genus Ammospiza. If many had been present on this date I would have seen them. On May 5 both species were plentiful on the Chadwick marshes. Furthermore, the Seaside Sparrows were defending territories. The absence of the two species the previous day indicates a large nocturnal flight.
It was during the second and third weeks in May that the sparrows of this genus were most abundant. In this period many unbanded Seaside Sparrows were in the patches of cattails that were being defended by the resident males from other territory-holders.
One Ammospiza caudacuta subvirgata (G. E. W. 545) was taken in the course of the study. It was a female (ovary: 7 x 5 mm.) weighing 15.3 grams ("moderate fat"), taken on June 8, 1955, on a marsh near the mouth of the Manasquan River on the Monmouth-Ocean County line. This marsh is decidedly less brackish than the Lavallette and Chadwick marshes. The specimen was the only Ammospiza seen there and was probably a migrant, despite the late date; this subspecies is known to occur late along the Atlantic Coast south of its breeding range. Cruickshank (1942:454-455) considers the peak of spring migration for this subspecies to be reached in late May.
To find A. c. subvirgata in a marsh seemingly not saline enough for the nominate race is not surprising. A. c. subvirgata breeds in marshes, along the coast of New England, which are almost fresh water (Montagna, 1942b:256). A. c. caudacuta is only casual away from salt water.
TERRITORY
In a general treatise on the subject of territorialism, Nice (1933:98), summarizing Howard, stated: "Territory implies in the male bird isolation, advertisement, fixation, and intolerance." I concluded from my observations that all four requirements are exhibited by male Seaside Sparrows while none of them is well developed in male Sharp-tailed Sparrows. This subject is discussed separately for the two species.
Tomkins (1941:38-51) studied populations of Ammospiza maritima macgillivrayii near the mouth of the Savannah River in South Carolina and Georgia and concluded that this form is not territorial. In support of his conclusions, he quoted Nice (1933:90-91) as follows: "Territory cannot mean just the nest spot when the adults feed in common; this may be 'nest territory,' but it is a very different matter from a territory in its strict sense to which parents confine themselves during the breeding season. Again, the very essence of a territory lies in its exclusiveness; if a bird's range is not defended, it is not a territory."
The feeding and nesting grounds of breeding Seaside Sparrows are often separated by a portion of the marsh which is not used by the birds. This complicates study of the territorial habits of the species. It does not mean, however, that the species is not territorial.
The birds studied by Tomkins had separate feeding and nesting grounds. Concerning this, Tomkins (1941:43) states that "The Seaside Sparrows of this locality [Savannah River area] often live where the two requirements [adequate feeding grounds and suitable nesting cover] are not always together or even meeting, but also where the feeding grounds and the nesting place are separated by a short distance."
Six of the eight original pairs of breeding Seaside Sparrows of the Lavallette colony fed in areas separate from those in which they nested. I found the eight nests of the original residents and banded and dyed all of the adults. The owners of two nests did not have separate nesting and feeding areas. One nest was built within fifteen feet of the south shore of the island, adjacent to the feeding area. The other was built within a few feet of the north shore. The female of this nest obtained food along the shore in the immediate vicinity of the nest. Her mate was absent; in all probability it was the singing male which I took from a nearby bush, before I found it advantageous to use the island as a study area.
The remaining six pairs flew to the south or west shores of the island in order to feed. None of these six nests was more than 100 yards from the feeding grounds (pl. 1, fig. b).
It was comparatively easy to see that the males defended an area surrounding the nest. It was more difficult to see that the pairs fed on separate plots of shoreline, each defended by the male, but I am convinced that this was the case.
The nest area was defended by the males through singing and chasing. I saw no instances of a female entering into territorial disputes; nevertheless, I did see a female chase a Sharp-tailed Sparrow away from the vicinity of her nest.
Tomkins (1941:46) did not consider the song of A. m. macgillivrayii to be "a declaration warning other birds away." After observing the behavior of males of A. m. maritima I am convinced that advertisement of intolerance is the primary purpose of song in this species. An account of the activities of a male Seaside Sparrow on May 6 on the marsh at Chadwick demonstrates this point. In an hour (6:01-7:01 a.m.) the bird sang 395 times, an average of 6.6 times per minute. He faced his nearest singing competitor when singing, which in the course of this hour was usually a male approximately 250 feet away across a creek. The two competitors almost always alternated their songs and frequently the singing of one seemed to stimulate the other bird to sing. Although the song of the Seaside Sparrow is short and unmusical it is loud, especially when compared with the song of the Sharp-tailed Sparrow. Elevated perches such as the tallest cattail stems or isolated bushes were used as singing and observation perches.
The chase of the Seaside Sparrow is not vigorous, but in all cases the intruder was seen to give way to the defender. I saw no physical fights between Seaside Sparrows. Chase by a defending bird was close to the ground and directly toward the intruder. Sometimes the attacking male emitted chipping notes when first sighting or flying towards his adversary.
In the hour of observation mentioned above, no other Seaside Sparrows entered the bird's territory, which consisted of a strip of cattail and shoreline, 250 feet long and no more than 25 feet wide. At other times Seaside Sparrows did enter this male's territory, and he drove them out as soon as he saw them. Savannah and Swamp Sparrows, which for a few weeks migrated through the area, were not chased, but Sharp-tailed Sparrows were chased at times.
Several times I flushed a particular male Seaside Sparrow from the northwest tip of the Lavallette study island so that it flew to the island to the north. Seaside Sparrows of this north island immediately made themselves conspicuous by chipping and then drove the non-resident individual back to its own territory.
The first time I heard what is described below as the social call of the Seaside Sparrow was on June 30 when an unbanded sparrow alighted in a marsh-elder bush near a nest. The individual called twice as it came near. The sound immediately aroused the owners of the nest and the male flew directly toward the strange bird. The intruder quickly and silently flew away.
My field notes refer to many other instances of territorial defense of the nesting area; it seems superfluous to cite them here.
Additional proof of territorialism in Seaside Sparrows was obtained by identifying and plotting the location of all the marked individuals, which I saw each day while systematically traversing all the available habitat on the island. Surprisingly, I did not once record a resident Seaside Sparrow in what I considered another male's territory in the month and a half (June 15-August 1) that I worked on the island at Lavallette.
The fact that the adult Seaside Sparrows did not search for food communally, or that different pairs did not utilize one particular area at different times was most apparent when the pairs were feeding young. From the blinds I first noted that the adults from any given nesting territory always flew in the same direction towards the shore. After moving a blind closer to the shore I noted that once an adult arrived at the open or sparsely vegetated shoreline, that adult restricted itself to a certain portion of the shoreline. These shoreline territories were plotted on field maps and appear on the map in plate 1, figure b. One method used to ascertain the boundaries of these shoreline feeding territories was a census taken from a boat. Many times I circled the island in a skiff identifying the marked sparrows as they appeared along the shore. The feeding sparrows were always found in the same areas around the island. Straight lines can be drawn between the nest sites and feeding areas of each pair of Seaside Sparrows without having any lines cross (pl. 1, fig. b). These lines correspond to the flyways used by each pair to go to and return from the feeding area. I consider the area defended about the nest, the segment of shoreline used by a pair of Seaside Sparrows and the connecting flyway to constitute the territory of a male Seaside Sparrow. If the flyways of any of the pairs had crossed, a situation of mutual exclusiveness would not have existed and a territory could not have been defined for the species.
It is generally agreed that territorial species engage in a minimum of fighting. Song and display have been evolved to substitute for actual combat which demands a greater amount of energy. Additionally, the mere presence of an individual in an area previously established as its territory probably serves to keep birds of nearby territories away. I think that male Seaside Sparrows defend the feeding area and flyway as a part of their territory by advertisement through use of these areas. The birds at Lavallette rarely sang on the feeding grounds and I noted only a few chases originating there. The sparrows rarely landed along the flyways. The constant use of these areas probably served as territorial defense, however. This supposition is supported by the fact that feeding areas and flyways of different pairs were mutually exclusive.
Nice (1941:457) divided territory into six categories. Type A (mating, nesting, and feeding ground for young) is the type exhibited by the Seaside Sparrow. The territory of a male Seaside Sparrow must contain an area of open mud and/or sparse vegetation where food can be obtained and also enough suitable cover to conceal the nest. I suspect in the case of the few males studied on the marshes at Chadwick that the territories the males established (strips of cattails and adjacent shoreline) did not have suitable nesting cover, because these males were unmated on June 15 when I left this study area because of human interference. Suitable nesting cover and feeding areas were separated by short distances of unusable marsh for most of the sparrows on the Lavallette study area. This fact caused the adults to commute from one site to the other. Photographs of shoreline habitat suitable for feeding by Seaside Sparrows appear in plate 2.
The area defended about the nest tended to follow the rows of marsh-elder bushes (pl. 3, fig. a), probably because these bushes supplied suitable song and observation perches. The segments of shoreline used by each pair were less than 75 yards in length and scarcely 20 feet wide. I never recorded Seaside Sparrows foraging in the interior of the marsh.
Sharp-tailed Sparrows were more abundant than Seaside Sparrows on the marshes at Chadwick and Lavallette. Sharp-tailed Sparrows were the more difficult to net because of the peculiar organization of the colonies. This organization, described below, also made nests of that species the more difficult to find. Only intensive netting at both localities produced enough marked individuals for me to study the breeding behavior of the species.
At Chadwick, where I netted most of the 85 Sharp-tailed Sparrows that I banded, my efforts were concentrated on one segment of the marsh. Marking made it evident that the males were not territorial, although they did confine themselves to what might appropriately be called a breeding home range, the area to which an individual confines itself in the course of one nesting attempt. Observations of marked birds also indicated that there was considerable overlap of the breeding home ranges of individual males.
I recorded a few marked Sharp-tailed Sparrows often enough and over a long enough period (more than one month) to gain a good idea of the size of the breeding home range of the males, which I estimate to be three to four acres. This estimate was made at Chadwick, where large areas of suitable uniform habitat occur. Females are more secretive than males, but seem to restrict themselves to areas considerably smaller than those of the males. My observations of two females that were feeding young indicated that each female restricted herself to an area of less than one acre. Female Sharp-tailed Sparrows possibly are territorial, although I recorded no disputes that would substantiate this possibility.
If I am correct in my estimates of size of breeding home range in Sharp-tailed Sparrows (males, three to four acres; females, approximately one acre), certain observations made by Montagna and me are readily explainable.
My netting operations indicated a surplus of male Sharp-tailed Sparrows in a given area. At Chadwick, I netted as many Sharp-tailed Sparrows as I could, without regard to sex. Here I captured 39 males and 16 females (six individuals remained unsexed). On the Lavallette study island, netting was more selective; here I attempted to net the females of the nests I found. The sex ratio at Lavallette was 15 males to eight females (one juvenile was not sexed). Three of the eight females were netted at their nests.
Montagna (1940:196) decided from collecting and observations that male Sharp-tailed Sparrows either outnumbered the females or were polygamous. The results I obtained from netting seemed to indicate a surplus of males. Banding, however, showed that in the breeding season males range over a larger area than do females. With this knowledge, the discrepancy between the number of males and females captured is explainable without an unbalanced sex ratio. If the males range over an area four times as large as that of the females, theoretically, four times as many males should be caught at every placement of the net provided the net remained in place long enough to capture all the birds using the area. In practice, this is essentially what occurred.
Other behaviorisms of this species indicate that it is not territorial. The song of the male is not loud and does not seem to be an advertisement to other birds. In fact, the song of this species is so quiet and lengthy when compared to that of the Seaside Sparrow that I at first thought I was hearing "whisper" or "practice" songs. These qualities of the song seem to indicate that the "advertising" function of song of territorial species is lacking or unimportant in Sharp-tailed Sparrows.
I suspect that male Sharp-tailed Sparrows do not even know where nests are. On July 18 at 7:00 a.m. I was watching a nest from a nearby blind when an unbanded male (I saw the individual sing later) appeared. As the bird foraged through the black grass, it headed directly toward the nest. When the male was almost one foot from the nest the incubating female left. She ran from the tussock and flew a short distance away to a cattail stem. From here she watched the male, which seemingly oblivious continued foraging, coming within inches of the nest. As the male walked away from the nest the female returned. At 8:00 p.m. the same day I was in the blind again. The female was out searching for food when a different, banded male appeared. In his foraging, the male walked up on the grass stems over the nest. The male apparently saw the young (two had hatched on July 17 and one on July 18) for he turned his head and seemed to peer down under the stems. The female appeared (with food) as he was doing this; she flew directly toward him and he flew away. The male was not seen near the nest in later observations.
On July 1 (6:50 a.m.) I was in a blind near another nest as the female approached with food for the young. At this moment a male appeared and the female immediately flew away. The male perched on a tussock within two feet of the nest, sang, and then flew off. The female reappeared in a few seconds without the food. She searched through a clump of black grass four feet from the nest, caught a small, pale green insect, fed it to her one young (there were also two eggs in the nest) and began brooding.
VOICE
Song
Only males of the two species sing. The normal song of the Seaside Sparrow lasts just under two seconds, the buzzing final note constituting three-quarters of the song. Saunders (1951:257-258) describes this song as short, and buzzlike, beginning with two or three short, rather faint notes and ending in a trill at first loud but fading away toward the end. The introductory notes are followed by a higher-pitched, loud, strongly accented, but buzzy note. This note is usually higher than the final trill and connected with it. The song has been written as tup tup ZEE reeeeeeeeee and tup TEE tle reeeeeeeeeee (Saunders, 1951:257), cutcut, zhé-eeeeeeeee (Peterson, 1947:232) and che-zheéeege, che-zhée, che-wéege, chur-zhée and too-szheée (Stone, 1937:910). My field notes contain the following: CHUR-er eeeee, CHUR eeeeee and oka-CHE weeeee. These variations in the phonetic representation of the songs are attributable mostly to the birds. Not only is there variation among individuals, but also individuals vary their songs. Birds that I heard giving a characteristic song suddenly sang a different type for awhile, and then reverted to the original. The bill is elevated and opens considerably with each note; the head bobs with the loud note. Typical singing postures are shown by Tomkins (1941: pl. 3).
The song of the Sharp-tailed Sparrow, as described by Saunders (1951:256-257), is short and insectlike, introduced by one or two short notes; the remainder is a somewhat wheezy trill, growing fainter towards the end. Sometimes there are two trills on different pitches, and occasionally a final short, low note. The quality is as though the sound sh ran through all but the introductory notes. Saunders writes these trills as: tsup tsup shreeeeeeeee and tip tish eeeeee shaaaaaaay. The bill is opened slightly with each note, as I saw when I watched a singing bird with the sun directly behind it. Montagna (1942a:116) noted that A. c. caudacuta sang less often than the more northern A. c. subvirgata.
Both species have specialized flight songs, but in the birds that I studied these songs were infrequent and seemingly unimportant. The flight song of the Seaside Sparrows consisted of a double version of the normal song. Although I heard it only a few times, the flight song of the Sharp-tailed Sparrow seemed slightly louder than the normal song. This song is given by both species as the bird flutters upward ten or 20 feet and glides back down.
Singing begins at daylight and decreases at 9 or 10:00 a.m. when the temperature rises. On cloudy days singing seemed to last longer. Towards dusk singing again increases, but not to the frequency of the morning peak.
The major differences between the songs of the two species are in loudness, length, and frequency. The fact that the Seaside Sparrow sings louder than the Sharp-tailed Sparrow is mentioned by Stone (1937:906). On windless days I heard singing Seaside Sparrows more than 200 yards away; Sharp-tailed Sparrows were inaudible at distances of more than 40 yards. The song of a Seaside Sparrow is rarely longer than two seconds; the song of a Sharp-tailed Sparrow usually lasts for almost 20 seconds and consists of a variable number of phrases like those described by Saunders. A Seaside Sparrow that I watched for one hour sang 395 times or 6.6 times per minute. I doubt that any of the Sharp-tailed Sparrows sang more than 20 times per hour, although I made no comparable count.
Additionally, Seaside Sparrows sing from exposed perches such as tall cattail stems and tall or isolated marsh-elder bushes. Sharp-tailed Sparrows do not often use conspicuous perches for singing. They sing while on the ground or when in flight. They do use exposed perches as observation posts and occasionally sing from them.
Seaside Sparrows often face their nearest neighbor when singing and alternate songs with this bird. The one time Sharp-tailed Sparrows almost always sing is when they are involved in fighting. In such a case the several birds sing simultaneously.
Seaside Sparrows began singing the morning after their nocturnal arrival. For resident birds, singing is at its maximum at this time and is maintained at a high level throughout incubation. At hatching of the eggs, singing declines sharply; males then are busy aiding in care of the young. Males that have successfully reared a brood rarely sing after the young leave the nest.
Sharp-tailed Sparrows sang infrequently when they first arrived, and singing did not reach its peak until late May. By August singing had almost ceased in this species.
Song of the Seaside Sparrow functions importantly in the establishment and maintenance of its territory. Newly-arrived males sing vigorously. In the Sharp-tailed Sparrow I think song is merely an expression of sexual excitement because song does not reach maximum frequency until the females arrive and become receptive to the males.
Differences in song correspond to differences in territorial behavior. The distinct, loud song, sung often and from exposed perches, which is frequently alternated with that of the nearest competitor, is given by the Seaside Sparrow, a territorial species. The indistinct, quiet song, sung infrequently and often from unexposed places belongs to the Sharp-tailed Sparrow, a non-territorial species.
Calls
Seaside Sparrows give a soft, lisping call note, probably the one referred to by Saunders as a squeaky tseep (1951:258), that functions as a social call. When migrants were numerous on the marshes at Chadwick I heard this note often. At Lavallette I did not hear it until June 30 (work began there on June 16) and then it was from an unbanded, non-resident bird. In late July and in August the number of non-resident sparrows increased and the social call was heard often. I never heard a resident bird give this call. On December 29, 1955, on a marsh at the mouth of the Manasquan River on the Monmouth-Ocean County line, a group of wintering Seaside Sparrows frequently used this call. I do not know whether the Sharp-tailed Sparrow has a comparable call.
Both species emit alarm notes. Although variable, the Seaside Sparrow has two general types. One, recorded by me as a short chip or tick was given by both sexes whenever I was near a nest. The other type, a high, sharp tsip, is indicative of a higher degree of excitement. When I captured young already out of the nest, or when I investigated nests containing young old enough to depart, the adults gave this call. The tail is jerked downward each time this note is given.
The alarm call of the Sharp-tailed Sparrow is not so loud as that of the Seaside Sparrow and it is not given so often. I described it as a short tsick or tsuck. Females emitted such calls when I was at their nests or when male Sharp-tailed Sparrows came near their nests. Males may have a similar call, but I never recorded it. Montagna (1942a:116) remarks on the quietness of this species. This is especially evident when one compares Sharp-tailed Sparrows with Seaside Sparrows.
PLATE 1
Fig. a. An aerial photograph of the Lavallette study island. One inch equals approximately 375 feet. The area covered by sand has been extended since this photograph was taken. This is indicated in figure b of this plate.
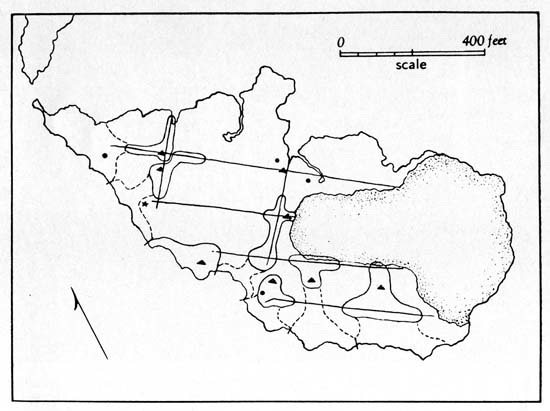
Fig. b. Map of the Lavallette study island. All fringillid nests that I found are indicated and the territorial boundaries of the Seaside Sparrows are shown.
▲—Seaside Sparrow
●—Sharp-tailed Sparrow
★—Song Sparrow
PLATE 2

Fig. a. The south shore of the Lavallette study island showing the two major driftlines and the sparsely vegetated areas. This is the feeding habitat of the Seaside Sparrow.

Fig. b. A close-up view of a segment of the shoreline. Note the spacing of the clumps of cord-grass (Spartina alterniflora). A six inch ruler propped against a 12 inch stick is included to indicate the size and spacing of the plants.
PLATE 3

Fig. a. The inner portion of the marsh on the Lavallette study island showing the rows of marsh-elder bushes (Iva frutescens) and the extensive areas of black grass (Juncus gerardi). Areas of mixed black grass and cord-grass appear in the foreground. All the nests of Sharp-tailed Sparrows were found in the areas of black grass. Four of the eight nests of Seaside Sparrows were in the marsh-elder. One of the blinds that I used is shown in this photograph.

Fig. b. A mated, banded pair of Seaside Sparrows in a dead marsh-elder bush near their nest. Note the abdomen of a moth protruding from the bill of the female on the right.
PLATE 4

Fig. a. A female Sharp-tailed Sparrow at the entrance to her nest. The throat on this bird is dark because of dye applied by me.

Fig. b. The nest of a Sharp-tailed Sparrow viewed from above. Stems of black grass were parted to take the picture. The outer rim of this nest (lower right) is made of living stems of black grass.
COPULATION
In late June at the Lavallette area there was an influx of unbanded Seaside Sparrows. Certain of these new arrivals established territories in areas unoccupied by the remaining original residents. These new residents were birds that probably had unsuccessful nestings elsewhere. Because of tropical storms that almost covered the island with water in August, I doubt that any of these late nestings were successful. On July 7 at 8:30 a.m., while watching a pair of these new arrivals, I recorded my only observation of copulation in the Seaside Sparrow. The female seemed to be searching for a nest site when copulation occurred. The female crawled about in a marsh-elder bush seemingly testing the various forks in the branches for size. The male followed her, remaining a few inches above and behind. Several times the two birds disappeared in the lower branches and were hidden by the surrounding black grass. Finally, while the female squatted on a branch the male mounted. He fluttered his wings before mounting and continued to do so as coition took place.
I began observations at Lavalette on June 16, too late to observe copulation of the early residents. All the nests contained eggs by that time. At Chadwick, pair formation seemingly never occurred, at least with the males I was studying. The territories established by males at Chadwick contained few marsh-elder bushes. Possibly females, finding no suitable nest sites, refused to accept these territories.
Copulation in the Sharp-tailed Sparrow was observed several times. It occurs most frequently in the course of, or immediately following, a fight between several males. I do not know what instigates the gathering of several males into these groups; it may be a certain behaviorism of a female, or possibly, merely the appearance of a female. Montagna (1942a:117) was convinced that females of A. c. subvirgata were present in these fights. On the other hand, in two instances with A. c. diversa where he collected all the birds in the group, no females were present. Twice, at Chadwick, my observations indicated that females of A. c. caudacuta were not always involved in these groups. In these instances all the birds in the group had previously been banded and diagnosed as males. Possibly a female was the original stimulus of these groups, and she may have disappeared while the males were fighting with each other. I found it difficult to distinguish fighting males from a copulating pair. On June 3, however, a banded pair was observed. Copulation occurred on the ground. The male fluttered his wings as he mounted and the female remained motionless. Copulation lasted approximately three seconds; immediately thereafter the male flew to a nearby cattail stem and the female climbed a tussock of grass and chipped quietly. This same female was seen to copulate with other males, and males were observed copulating with several females.
A. m. maritima is monogamous, the pair-bond being maintained throughout a breeding cycle. A. c. caudacuta is promiscuous, relations between the sexes being limited to copulation. For A. c. subvirgata a relationship other than promiscuity has been intimated (Lewis, 1920:587-589). Concerning observations of the nest he found at Yarmouth, Nova Scotia, Lewis wrote: "The nest was found after I had quietly watched the parent Sparrows for about an hour, while they were bringing food to their young.... The male sang from time to time from a piece of driftwood on the marsh about 30 feet distant from the nest. When I was examining the nest and the young birds, the parents made no demonstration for some minutes, but later they came near and uttered chip's, much like those of Savannah Sparrows."
NESTS
I found the nests of all eight pairs of Seaside Sparrows which nested on the Lavallette study island in 1955. Four nests were supported by marsh-elder bushes, three of which were dead. These nests were placed low enough to be hidden by numerous stems of black grass, as were the other four nests. Of the remaining four nests, three were placed in tussocks of black grass and the fourth one gained support mostly from cord-grass stems. The eight nests ranged from 9 to 11 inches (9.6 inch average) from the rim to the ground, the four nests in the bushes being the highest. The outside diameters of the nests ranged from 3 to 4.5 inches (3.9 inch average) and the outside depth varied between 2 and 3.5 inches (2.7 inch average). Seven of the nests had an inside depth of 1.5 inches; the other one was only an inch from the rim to the floor. The inside diameter of the cup varied between 2 and 2.5 inches.
As mentioned above all eight nests were shielded by stems of black grass. Stems were not woven over the nests by the birds; rather it was the choice of the nest sites that resulted in the concealment. The only plant used for nest material was black grass.
In all cases the black grass limited the directions from which the nests could be entered. Six of the nests were approached from a direction varying between northeast and southeast. The prevailing winds of spring and summer are from the south and southwest; the black grass consequently leans in the opposite direction. The remaining two nests were entered from the northwest. These were nests built in marsh-elder bushes where the grass stems were held upright by the branches of the bushes.
One nest, built in a small dead marsh-elder bush, was tilted by the growth of stems of black grass which were used for support on one side. This tilting did not cause the contents to spill, but, I judged, did cause the adults to desert the nest.
Seven nests of the Sharp-tailed Sparrow were found; two of these were old nests. Four of the five nests used in the breeding season of 1955 were found on the Lavallette marsh study area, the other one I discovered on the Chadwick marshes. Two young Sharp-tailed Sparrows that I saw at Lavallette were not from nests I found, nor were they from the same nest. Therefore, a minimum of six Sharp-tailed Sparrows bred on the Lavallette island. Measurements were taken of only the five nests that were used in 1955. The Sharp-tailed Sparrow builds its nest closer to the ground than does the Seaside Sparrow. The five nests were five to six inches off the ground; the two nests of a previous year appeared to have been no higher. The Sharp-tailed Sparrow nests were built in areas where black grass was the predominant plant, and the nests were constructed entirely from this grass. The outside diameters varied from 3 to 4.25 inches (3.4 inch average). The outside depth of the nests varied from 2 to 3.5 inches (2.8 inch average). The inside depth was 1.5 inches in all nests and the inside diameter ranged from 2 to 2.5 inches (2.1 inch average).
Harrison F. Lewis (1920:587) studied a nest of A. c. subvirgata in a small salt marsh near Bunker's Island at the southern end of Yarmouth Harbor, Yarmouth, Nova Scotia, which he found on June 12, 1920. For details of this nest I quote Dr. Lewis. "The nest proper was a neat, round cup of fine, dry, dead grass, with some horsehair in the lining. Its foundation consisted of some small masses of 'eel-grass' and roots. Its dimensions were: inside diameter, 2.5 in.; outside diameter, 4.5 in.; inside depth, 1.5 in.; outside depth 2.375 in. It was elevated above the general surface of the marsh by being placed on the top of a low, grassy ridge, about fourteen inches high, formed from material thrown up when a ditch was dug across the marsh, many years before. During some storm a mat of dead 'eel-grass' had been left on top of this ridge, and this had later been lifted by the growing marsh grass, leaving several inches between it and the ground. The nest was placed on the northwest edge of this mat, about half of the nest being under it, while the other side was sheltered and concealed by grass about six inches high. The nest was not sunk in the ground at all."
Two of the nests found were entered from the north-northeast, the other three from the east-southeast. All five nests were sheltered above by stems of black grass. Three of the nests were beneath a layer of dead black grass where a clump of erect living stems parted the mat. One nest (pl. 4, fig. a) was situated where cattail stubs held the black grass somewhat erect. Green stalks as well as dead stalks were woven into a canopy over this nest. Another nest was constructed on a mat of black grass under and among numerous horizontal living stems, some of which were woven into the outer lining of the nest (pl. 4, fig. b).
Nests of both species were found in tussocks of black grass. The locations of these sites differed. The Sharp-tailed Sparrow prefers the higher and therefore dryer portions of the marsh where black grass is the characteristic plant. Contrastingly the Seaside Sparrow almost always chooses the wetter portions of the marsh (Cruickshank, 1942:45; Forbush and May, 1939:514; Stone, 1937:906; personal observations) where several species of plants are abundant. In areas that have been ditched, as have almost all marshes in New Jersey, the mound of excavated muck is ideal for the growth of marsh-elder. Rows of these bushes are present on many of the marshes of New Jersey (pl. 3, fig. a). The location of four of the eight Seaside Sparrow nests in these "hedgerows" indicates that they provide suitable, if not preferred, sites for the species.
EGGS AND INCUBATION
I found no nests of either species before they contained a complete complement of eggs and therefore was unable to ascertain the incubation period for these species. Brood patches were evident on female Sharp-tailed Sparrows by June 1, probably indicating that laying began near this date. Cruickshank (1942:456) lists egg dates of the Sharp-tailed Sparrow as concentrated in early June, with extremes of May 19 and August 4. He thinks the species probably has two broods. For the Seaside Sparrow, Cruickshank (1942:458) states there is probably but one brood and that egg dates are concentrated in early June, with extremes May 23 and July 2. Stone (1937:907, 911) considers four eggs a normal clutch for both species, but cites instances where three and five eggs were thought to be complete sets. Four of the eight Seaside Sparrow nests I found contained at least three eggs, and four contained at least four eggs. Four of the five Sharp-tailed Sparrow nests I found contained at least three eggs and one contained four eggs.
Female Seaside Sparrows do all of the incubation. The male, while the female is on the nest, remains a short distance away. He sings often and gives alarm notes when there is a local disturbance. These chipping notes bring the female off the nest, and then they both chip at the intruder. The male accompanies the female to the feeding grounds and normally they return together.
As previously mentioned, male Sharp-tailed Sparrows take no part in the nesting activities.
YOUNG
I studied growth and changes in behavior of the young. Since I could see no behavioral differences between the nestlings of the two species, this subject will be discussed jointly for the two forms.
Growth
The color of the natal downs of both species is similar. Dwight (1900:190), who saw newly hatched nestlings only of the Sharp-tailed Sparrow, described the color as grayish wood-brown. A series of white neossoptiles is present at the posterior end of the ventral tract in both species. These feathers are more numerous in the Seaside Sparrow. Dwight (1900:98) saw no neossoptiles on the underparts of any of the passerines he examined. Seaside Sparrows have a mid-dorsal row of downs in the dorsal tract near the uropygium. These feathers are lacking in the Sharp-tailed Sparrow and constitute the major difference, in this plumage, between the two species. The neossoptiles of three Seaside Sparrows and one Sharp-tailed Sparrow were counted. These counts were checked on the young birds studied in the field. The number and placement of these feathers appear in plate five. There appears to be a consistently greater number of natal downs in Seaside Sparrows, when compared with Sharp-tailed Sparrows.
Table 1—Daily Weight in Grams of Nestling Seaside Sparrows and Sharp-tailed Sparrows from Lavallette, Ocean Co., New Jersey.
| Day | Ammospiza maritima | Average | ||||||
| 0 | ... | 2.2 | 2.3 | ... | ... | 2.2 | 1.8 | 2.1 |
| 1 | 3.0 | 3.3 | 3.5 | 3.7 | 3.3 | 2.6 | 2.1 | 3.1 |
| 2 | 4.6 | 5.0 | 5.2 | 5.0 | 4.8 | 3.7 | 3.0 | 4.5 |
| 3 | 7.0 | 5.5 | 7.2 | 6.9 | 6.9 | 5.9 | 4.7 | 6.3 |
| 4 | 9.4 | 8.1 | 10.6 | 9.1 | 9.1 | 7.6 | 6.4 | 8.6 |
| 5 | 12.5 | 11.1 | 12.3 | 11.4 | 11.1 | 9.9 | 8.7 | 11.0 |
| 6 | 14.6 | .1 | [1]11.1 | 13.9 | 13.7 | 12.6 | 9.6 | 13.0 |
| 7 | [1]11.6 | 13.9 | 12.1 | 15.1 | 14.8 | 14.3 | 11.8 | 13.7 |
| 8 | 14.9 | 15.5 | 13.4 | 14.9 | 14.8 | 14.6 | 12.4 | 14.4 |
| 9 | 15.2 | 15.8 | 13.8 | 16.2 | 16.1 | 16.0 | 14.4 | 15.4 |
| 10 | ... | ... | ... | ... | 15.9 | 15.5 | 14.3 | 15.2 |
| Day | Ammospiza caudacuta | Average | ||||
| 0 | 1.6 | ... | 1.8 | 1.7 | 1.7 | 1.7 |
| 1 | 1.8 | 2.3 | 2.5 | 3.0 | 2.1 | 2.3 |
| 2 | 3.3 | 2.7 | 4.1 | 4.6 | 3.1 | 3.6 |
| 3 | 5.0 | 3.7 | 5.9 | 6.4 | 4.7 | 5.1 |
| 4 | 6.8 | 5.4 | 8.4 | 9.1 | 6.7 | 7.3 |
| 5 | 8.6 | 6.9 | 10.7 | 11.2 | 9.5 | 9.4 |
| 6 | 10.2 | 8.9 | 12.8 | 13.0 | 10.9 | 11.2 |
| 7 | 12.1 | 11.4 | 14.5 | 13.6 | 12.3 | 12.8 |
| 8 | 13.5 | 12.9 | 15.3 | 14.5 | 13.3 | 13.9 |
| 9 | 12.2 | 13.4 | 15.9 | 14.9 | 13.6 | 14.0 |
| 10 | 12.7 | 14.0 | 15.5 | 15.0 | 14.0 | 14.2 |
| 11 | ... | ... | ... | 15.1 | 14.4 | ... |
[1] These weights are not figured in the averages; see text.
Seven nestling Seaside Sparrows and five nestling Sharp-tailed Sparrows were weighed at 24-hour intervals until they left their nests. The birds were weighed in early morning before they had received much food. Weights of these individuals, and daily averages for each species are shown in Table 1. The weights in the zero column were of nestlings that had not been fed. The weight of one hatchling (1.9 gm.), which does not appear in the table, is included in the average for the zero column. Two young Seaside Sparrows, approximately a week old, fell out of a nest between 9:30 a.m. July 6 and 5:30 a.m. July 7. When I found them below the nest, at the latter time, their temperatures were far below normal, and they had lost a considerable amount of weight. These abnormally low weights were not figured in the averages. The weights of Sharp-tailed Sparrows 11 days old were obtained by confining the birds to the vicinity of the nest with a screen.
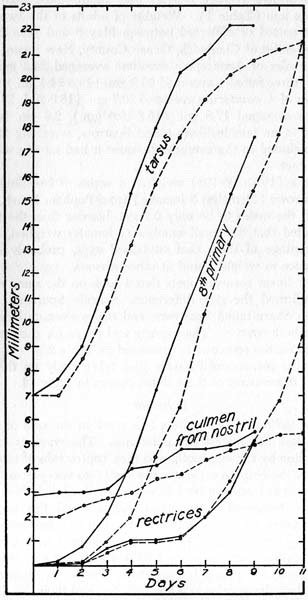
Fig. 1. The development of the young of Seaside Sparrows (solid line) and Sharp-tailed Sparrows (dotted line) as evidenced by four linear measurements taken at 24 hour intervals.
At hatching and throughout nestling life and post nestling life Seaside Sparrows average heavier than Sharp-tailed Sparrows of comparable age (Table 1). Weights of adults of the two species that were netted or collected between May 6 and June 27, 1955, within two miles of Chadwick, Ocean County, New Jersey, follow: Fourteen males of Ammospiza maritima averaged 24.2 gm. (21.9-27.4 gm.); three females averaged 22.3 gm. (19.8-24.4 gm.). Thirty-three males of A. caudacuta averaged 20.7 gm. (18.0-23.1, 25.8 gm.); 14 females averaged 17.8 gm. (15.3-19.0 gm.), 2.9 gm. less than the males. One female Sharp-tailed Sparrow, weighing 23.1 gm., was not included in the averages because it had an egg with shell in the oviduct.
Montagna (1940:195-196) weighed a series of breeding Sharp-tailed Sparrows (21 males; 5 females) from Popham Beach, Maine, and found the males to be only 0.2 gm. heavier than the females, but he stated that the small number of females weighed, and the high percentage of these that contained eggs, probably lessened the difference in weight found at other seasons.
The four linear measurements that I took on the same series of adults confirmed the size difference: Seaside Sparrows average larger than Sharp-tailed Sparrows, and males average larger than females in both species. The average and range for each measurement taken on the sparrows is presented in Table 2.
Four linear measurements were also taken daily on the young sparrows. A summary of these data appears in Figure 1.
Behavior
The first indication of hatching is a crack in the side of the egg along the line of greatest circumference. The crack is extended along this line by the egg tooth, and then contraction of muscles of the neck by the embryo separates the shell into two pieces. Extension of the legs frees the bird from the shell. I held the eggs of two Seaside Sparrows in my hand and watched this procedure. In each instance the young bird defecated in the shell before freeing itself. A barely audible "peep" note was heard from one hatchling Sharp-tailed Sparrow when I held it near my ear. When free from the shell, the young birds rest on their tarsi, abdomen and forehead; their down dries in a few minutes, and their skin becomes noticeably darker. One sparrow gaped five minutes after hatching and all the young gaped later the same day. The abdomen of the young becomes distended when they are fed by the parents.
PLATE 5
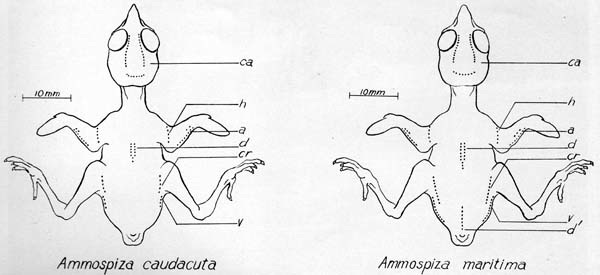
Ammospiza caudacuta Ammospiza maritima
Drawings of the nestlings of the two species of Ammospiza approximately three days of age showing the variation in the amount and placement of the neossoptiles in the two species. Abbreviations for feather tracts in which downs were found: ca, capital; h, humeral; a, alar; d, d´, dorsal; cr, crural; v, ventral.
PLATE 6

An aerial view of the marshes at Chadwick (upper left) and Lavallette (lower left). The Atlantic Ocean appears in the upper right of this photograph.
Table 2—Linear Measurements in Millimeters of Adult Seaside Sparrows and Sharp-tailed Sparrows Captured or Collected Within Two Miles of Chadwick, Ocean County, New Jersey, Between May 6 and June 27, 1955.
| Ammospiza maritima | ||
| 14 males | 3 females | |
| wing (chord) | 64.14 (60-66) | 58.33 (58-59) |
| tail | 55.28 (54-59) | 51.00 (49-53) |
| tarsus | 23.00 (22-25) | 22.17 (21-23) |
| culmen | 15.18 (15-15.5) | 14.50 (13.5-15.0) |
| Ammospiza caudacuta | ||
| 33 males | 15 females | |
| wing (chord) | 58.79 (55-61) | 55.67 (54-58) |
| tail | 49.48 (46-53) | 46.93 (45-50) |
| tarsus | 20.91 (20-22) | 20.30 (20-21) |
| culmen | 13.67 (13-14) | 13.23 (12.5-14.0) |
In the first 24-hour period after hatching the soft "peep" note is heard frequently. The young are better able to right themselves, and many feather papillae show distinctly through the skin.
On the second day young are capable of moving short distances by using their wings and feet. A thick ridge of tissue forms over the eyeball where the eyelids later delaminate. The call is now a double version of the "peep" note described above.
When the young are three days old the eyelids open, but only slightly. In the next three days the young become better co-ordinated and the eyes open fully. The egg tooth was last seen on a young bird on the sixth day. All incoming feathers remain sheathed until the seventh day.
On the seventh day young show the first signs of cowering. Previously, they all begged when I came to the nest. The remiges remain sheathed, but the body feathers emerge from the tips of the sheaths. A quiet reedy call replaces the "peep" note. A quiet, but squealing distress call was also first noted on the seventh day, when the young were handled.
On the eighth day the remigial sheaths become gray (previously they were dark blue) and begin to slough off. When removed from the nest, the young attempt to escape. Begging is less frequent and cowering is the predominant attitude towards intruders.
The first young of both species left the nest on the ninth day. It must be remembered, however, that this remark, and succeeding remarks, concerning departure of young from nests pertains to young that were disturbed daily by me. The others climbed to the edge of the nest when they were left alone, but remained in the nest when they were all replaced. Gaping was recorded once on the ninth day. Stuart W. John watched two Sharp-tailed Sparrows on my study area leave a nest. They climbed out and immediately hid in a tussock of grass a few inches behind the nest.
On the tenth day when I parted the branches over one Seaside Sparrow nest, the four young jumped from the nest and scattered in the grass. One of these birds gave a chipping note similar to the distress call of adults. No bird remained in a nest longer than ten days. Four young left the nest after nine days, seven young left on the tenth day. When the young leave the nest they are able to run rapidly through the dense grass. The young are fed by the parents for approximately 20 days after they leave the nest. Twenty-three days after one young Seaside Sparrow left the nest it was netted at the opposite end of the island, 300 yards from the territory of its parents.
Young Seaside Sparrows fly in an uncertain but characteristic manner when they are flushed from the grass. They dive clumsily into the grass after a short flight, making it easy to identify them as birds of the year.
The plumage of sparrows of the Genus Ammospiza serves to conceal them in their habitat. In juvenal and adult plumage, the Sharp-tailed Sparrow is a brown-backed, streaked bird, the color and pattern blending with the matted grasses (Allen, 1925:67) where the species feeds and nests. The Seaside Sparrow, as an adult, is olive-gray. Its color corresponds to that of the substratum where the species forages. The juvenal plumage of the Seaside Sparrow resembles that of the Sharp-tailed Sparrow. I believe that young Seaside Sparrows have this brown, streaked plumage because they spend most of their time in the dense grass. In the Seaside Sparrow a complete post-juvenal molt begins in late August. The resulting plumage resembles that which is acquired by the adults when they complete their post-nuptial molt (Dwight, 1900:192-193).
FOOD, FEEDING, AND CARE OF THE YOUNG
The food habits of Seaside Sparrows and Sharp-tailed Sparrows have been studied by Judd (1901:64-66), who concluded that both species are highly insectivorous. In 51 stomachs of Sharp-tailed Sparrows 81 per cent of the contents was animal. The results of investigation of stomachs of Seaside Sparrows were similar. In each of the two species the bill is more elongated and less conical than in other sparrows. For the two species studied, the shape of the bill seems to be an adaptation for feeding on insects.
When searching for food, Sharp-tailed Sparrows walk through the dense black grass, deftly brushing stems aside with their bill as they go. Open areas are generally traversed by rapid running. I never noticed either species hopping. They stop to investigate openings in the matted understory of grass, often sticking their heads into the holes. Many times I saw these sparrows stretch or jump to pick insects from stems. Many droppings, almost certainly those of Sharp-tailed Sparrows, were present in areas of damp, matted grass. Females, when feeding young, obtain most of the food near the nest; several times I saw birds catch insects when they were within inches of their nest. Sharp-tailed Sparrows feed also along the banks of pools and creeks, and along the perimeters of marshes. Sharp-tailed Sparrows seem to be less restricted in the types of feeding habitats they can use than are Seaside Sparrows.
Seaside Sparrows always returned to the edge of the marsh to procure food, according to my observations. The birds at Lavallette fed extensively on noctuid moths. In the feeding territories of two pairs of Seaside Sparrows, along the strip of washed-up eel grass, I found at least 40 wings of these moths. In several instances the four wings of one moth were lying close together in the same relative position in which they had been on the animal. Legs and pieces of thorax were also discarded occasionally. I watched adults take these moths from the stems of the smooth cord-grass and snip the wings off with their bills. Moth wings were present in the other feeding territories, but not in so great a quantity. Once I saw a female return to the nest with a spider in her bill. Spiders were abundant throughout the marsh.
Dwight (1900:193) was surprised that the two species living in the same environment, and therefore suffering equally from abrasion from the coarse marsh grasses, should have a different number of molts per year. The Sharp-tailed Sparrow has a complete pre-nuptial, as well as a complete postnuptial, molt. The Seaside Sparrow has only a postnuptial molt, the nuptial plumage being acquired by wear. My observations of the feeding habits of the two species indicate that they do not live in precisely the same environment. The Sharp-tailed Sparrow, which has two complete molts annually, generally forages in dense, abrasive vegetation. The Seaside Sparrow, which has but one molt each year, forages in relatively open areas.
Several times I saw adult Seaside Sparrows fly from their nests toward the feeding territories with fecal sacs in their bills. On the feeding grounds, I found several of these sacs discarded near the moth wings. I saw also female Sharp-tailed Sparrows leave their nests with fecal sacs. I did not see sparrows of either species swallow fecal sacs.
One nest, that of a Seaside Sparrow containing four young, became fouled with excrement when the young were nine to ten days old. It is interesting that these young were cared for only by a male, at least for the last four days of nest life, and that one of the young birds died two days before the others left the nest. This male's mate was probably a female that I banded on June 18 (the young left the nest on June 23) and never saw again. A female, whose mate was probably killed by me on June 15, continued to incubate the three eggs until they hatched on June 29, but deserted the nest when the young were two days old. This female was seen again on August 1 more than 500 yards from her nest site on the island immediately north of the study area.
Devotion of parent passerine birds to the young typically increases with the growth of the young (Nice, 1943:245). This may explain why the mateless female deserted its nest when the young were only two days old, whereas a mateless male continued to care for his six-day-old young. The death of one nestling, and the eventual fouling of his nest may indicate that the job was too much for one adult to perform. The correlation of increasing devotion of the parents with increasing age of the young was further illustrated by the distraction display, noted by me, on the part of a pair of Seaside Sparrows on the day their young left the nest. As I lifted the four nine-day-old young from the nest for weighing, they began to give the distress call. This attracted the parents from the feeding area approximately 60 yards away. The two adults ran around on the ground within ten feet of me giving the tsip note and fluttering their wings. Several times the adults flew within a few feet of me, making a vibrating sound with their wings. Although I realized the function of this display, it was distracting nevertheless.
ACKNOWLEDGMENTS
I am indebted to Assistant Professor Harrison B. Tordoff for comments and suggestions throughout the preparation of this manuscript, and to Mr. Stuart W. John who photographed the birds and the habitat scenes. The aerial photograph of the study island is reproduced with the permission of Fairchild Aerial Surveys, Inc., and the photograph of Lavallette and Chadwick through the kindness of the Lavallette Yacht Club. The drawings were made by Mr. John R. Beeder. Additionally I wish to express my gratitude to Dr. Dean Amadon, Mr. H. Lyman Sindle, and Mr. Lester B. Woolfenden for help and advice in certain aspects of the field work.
SUMMARY
A comparative study of the breeding behavior of the Seaside Sparrow and Sharp-tailed Sparrow was made in New Jersey in 1955.
Observations of marked individuals indicate that the Seaside Sparrow is monogamous and territorial, whereas the Sharp-tailed Sparrow is promiscuous, and at least the male is non-territorial. The male Seaside Sparrow defends its territory by chasing and singing. The male Sharp-tailed Sparrow confines itself to a breeding home range. This range is not a territory; it is inhabited by several males. Female Sharp-tailed Sparrows may be territorial; this is not certainly known.
The Seaside Sparrow sings louder, more distinctly, more often, and from more exposed perches than does the Sharp-tailed Sparrow. These characteristics seem to be correlated with territorial habits. Other calls are described and their functions are discussed.
The Seaside Sparrow nests in marsh-elder bushes, or in areas of mixed vegetation. The Sharp-tailed Sparrow prefers the inner, drier areas of a marsh, where black grass is dominant. The Seaside Sparrow places its nest farther above the ground than does the Sharp-tailed Sparrow. Both species used only black grass in constructing the nest.
Copulation is described. The incubation period was not determined for either species. Three or four eggs seem to be a normal clutch. Females do all of the incubating.
The young remained in the nests nine to ten days. These nests, of course were disturbed, for I visited them at least daily. The nestlings of the Seaside Sparrow are fed by both parents. Male Sharp-tailed Sparrows seem not to know the location of the nests and take no part in rearing the young at least up to time of fledging. The natal down of both species is described. Data on growth and behavior of the young are presented.
Seaside Sparrows obtained most of their food from the shoreline of the marsh, in areas of open mud and smooth cord-grass. The plumage of the adult matches, in color, this mud. The Sharp-tailed Sparrow feeds everywhere in the marsh, but mostly in areas of dense and matted black grass. The plumage on the dorsum of this species is brown and streaked resembling the dead grass. Juvenal Seaside Sparrows, which spend most of their time concealed in the dense grass, resemble adult and juvenal Sharp-tailed Sparrows in plumage. Sharp-tailed Sparrows molt completely twice per year. The Seaside Sparrow molts but once per year. The difference in number of molts, too, is correlated with habitat preference, since the grassy forage habitat of the Sharp-tailed Sparrow must result in greater abrasion of the plumage than does the open feeding habitat of the Seaside Sparrow.
LITERATURE CITED
| Allen, Glover M. | |
| 1925. | Birds and their attributes. Marshall Jones Co., Boston, Massachusetts. xiii + 338 pp., frontispiece, 45 pls., 6 figs. |
| Cruickshank, Allan D. | |
| 1942. | Birds around New York City, where and when to find them. The American Museum of Natural History, Handbook Series, No. 13, New York. xvii + 489 pp., frontispiece, 35 pls. |
| Dwight, Jonathan | |
| 1900. | The sequence of plumages and moults of the passerine birds of New York. Annals N.Y. Acad. Sci., Vol. XIII, pp. 73-360, 7 pls. |
| Forbush, Edward Howe and May, John Bichard | |
| 1939. | Natural history of the birds of eastern and central North America. Riverside Press, Cambridge, Massachusetts. xxv + 553 pp., 97 pls. |
| Griscom, Ludlow | |
| 1944. | A second revision of the Seaside Sparrows. Occ. papers of the Museum of Zool., L.S.U., No. 19. |
| Judd, Sylvester D. | |
| 1901. | The relation of sparrows to agriculture. U.S.D.A. Bull. No. 15, 98 pp., 4 pls., 19 figs. |
| Lewis, Harrison F. | |
| 1920. | Notes on the Acadian Sharp-tailed Sparrow (Passerherbulus nelsoni subvirgata). Auk, 37:587. |
| Montagna, William | |
| 1940. | The Acadian Sharp-tailed Sparrows of Popham Beach, Maine. Wilson Bull., 52:191-197, 2 figs., 1 table. |
| 1942a. | The Sharp-tailed Sparrows of the Atlantic coast. Wilson Bull., 54:107-120, 4 figs. |
| 1942b. | Additional notes on Atlantic coast Sharp-tailed Sparrows. Wilson Bull., 54:256. |
| Nice, Margaret Morse | |
| 1933. | The theory of territorialism and its development. Fifty years' progress of American ornithology 1883-1933. A.O.U., Lancaster, Pennsylvania, pp. 89-100. |
| 1941. | The role of territory in bird life. Amer. Mid. Nat., 26:441-487. |
| 1943. | Studies in the life history of the Song Sparrow II. Trans. Linn. Soc. N.Y. Vol. VI, viii + 329 pp., frontispiece, 6 figs. |
| Peterson, Roger Tory | |
| 1947. | A field guide to the birds. Houghton Mifflin Co., Boston, Massachusetts. xxiv + 290 pp., 60 pls. |
| Salt, W. Ray | |
| 1954. | The structure of the cloacal protuberance of the Vesper Sparrow (Pooecetes gramineus) and certain other passerine birds. Auk, 71:64-73, 5 figs. |
| Saunders, Aretas A. | |
| 1951. | A guide to bird songs. Doubleday and Co., Garden City, New York. xiv + 307 pp., 201 figs. |
| Stone, Witmer | |
| 1937. | Bird studies at Old Cape May. D.V.O.C. Philadelphia, Vol. II, pp. 521-941. |
| Tomkins, Ivan R. | |
| 1941. | Notes on Macgillivray's Seaside Sparrow. Auk, 58:38-51, pls. 2, 3. |
Transmitted June 14, 1956.
End of the Project Gutenberg EBook of Comparative Breeding Behavior of
Ammospiza caudacuta and A. marit, by Glen E. Woolfenden
*** END OF THIS PROJECT GUTENBERG EBOOK COMPARATIVE BREEDING ***
***** This file should be named 36285-h.htm or 36285-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/3/6/2/8/36285/
Produced by Chris Curnow, Joseph Cooper and the Online
Distributed Proofreading Team at http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
http://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.