
(1) CaSO3 + 2HCl = CaCl2 + SO2 + H2O
(2) Na2SO3 + 2HCl = 2NaCl + SO2 + H2O
The Project Gutenberg EBook of The Puering, Bating & Drenching of Skins, by Joseph Turney Wood This eBook is for the use of anyone anywhere in the United States and most other parts of the world at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org. If you are not located in the United States, you'll have to check the laws of the country where you are located before using this ebook. Title: The Puering, Bating & Drenching of Skins Author: Joseph Turney Wood Release Date: October 26, 2018 [EBook #58168] Language: English Character set encoding: UTF-8 *** START OF THIS PROJECT GUTENBERG EBOOK PUERING, BATING, DRENCHING OF SKINS *** Produced by Thiers Halliwell, Chris Curnow and the Online Distributed Proofreading Team at http://www.pgdp.net (This file was produced from images generously made available by The Internet Archive)
In this e-book a black underline indicates a hyperlink to a page, illustration or footnote (hyperlinks are also highlighted when the mouse pointer hovers over them), and a red dashed underline indicates the presence of a concealed comment which can be viewed by hovering the mouse pointer over the underlined text. Page numbers are shown in the right margin and footnotes are at the end. Footnotes are listed at the end.
The text contains tables, equations and chemical formulae that might not display correctly on small reading devices – a standard web browser is recommended.
The text of the book has mostly been preserved in its original form, including inconsistent italics, capitalisation, and hyphenation. However, some defects have been corrected silently, such as missing/inconsistent punctuation, parentheses and spacing, and an erroneous ÷ has been replaced by − in a fraction in footnote 69. Tables have been formatted to match the originals, but a large table that was originally split over four pages (pp. 98–101), has been condensed into two sections. Some illustrations have been positioned closer to the relevant text and the location may not correspond to that specified in the list of illustrations.
The following typographic corrections have been made:
mutiplied → multiplied
Kothbeize → Kothbeizen
of → on
phenolphtalein → phenolphthalein
hydroxylion → hydroxyl ion
Ca(OH2) → Ca(OH)2
pepsine → pepsin
bibliographic xref (9) → (19)
The following inconsistent spellings have not been altered:
Frankfort a/M – Frankfort o/M
Zenckeri – zenkeri
Casenin – Caseuin
Kentniss – Kenntnis – Kenntniss
resistant – resistent
c.c. – c.cm.
ureaæ – ureæ
ae – æ
The cover image of the book was created by the transcriber and is placed in the public domain.
THE
PUERING, BATING & DRENCHING
OF SKINS
Scientia et potentia humana in idem coincidunt, quia ignoratio causæ destituit effectum. Natura enim non nisi parendo vincitur: et quod in contemplatione instar causæ est, id in operatione instar regulæ est.
Knowledge and human power are synonymous, since the ignorance of the cause frustrates the effect; for nature is only subdued by submission, and that, which in contemplative philosophy corresponds with the cause, in practical science becomes the rule.—Bacon, Aphorism III.
DEDICATED
TO
SIR JOHN TURNEY
THE VETERAN
OF THE
LIGHT LEATHER INDUSTRY
IN
GRATEFUL ACKNOWLEDGMENT
| PAGE | ||
| LIST OF ILLUSTRATIONS | ||
| PREFACE | ||
| I. | DESCRIPTION OF THE PUERING AND BATING PROCESS | |
| II. | THE CHEMISTRY OF BATING | |
| III. | THE PHYSICS OF BATING | |
| IV. | THE BACTERIOLOGY OF THE BATE | |
| V. | ACTION OF ENZYMES | |
| VI. | ORIGINAL PAPERS ON BATING | |
| VII. | ARTIFICIAL BATES | |
| VIII. | PATENTS | |
| IX. | DRENCHING | |
| X. | ORIGINAL PAPERS ON DRENCHING | |
| XI. | BIBLIOGRAPHY | |
| CONCLUSION | ||
| INDEX |
| FIG. | PAGE | |
| 1. | Curve showing removal of lime by washing | |
| 2. | Cubical “truck” for measuring skin | facing 8 |
| 3. | Puer wheel | " 13 |
| 4. | Front elevation of puer wheel | |
| 5. | End elevation and section of puer wheel | |
| 6. | Sir John Turney’s scudding machine | facing 17 |
| 7. | Curves of ash contents during puering | |
| 8. | Diagram of volumenometer | |
| 9. | " connexions of electrometric apparatus | |
| 10. | Electrometric apparatus | |
| 11. | Apparatus for measuring degree of falling | |
| 12. | Curves obtained by measuring apparatus | |
| 13. | Improved apparatus for measuring degree of falling | |
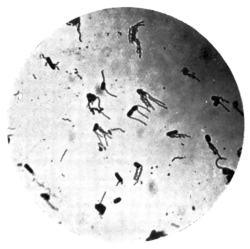
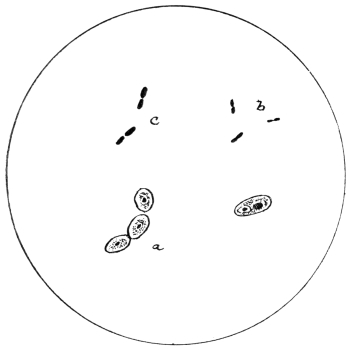
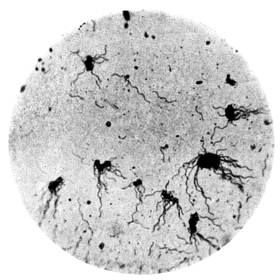
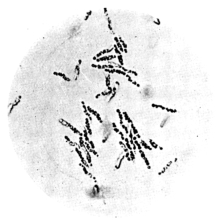
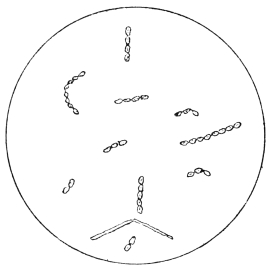
| 14. | Various forms of bacteria in puer liquors | facing 88 |
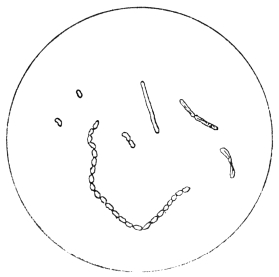
| 15. | B. coli commune | " 88 |
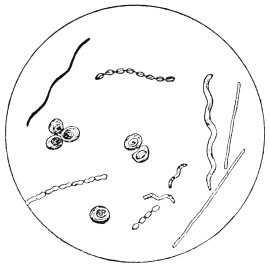
| 16. | B. erodiens (Becker) | " 97 |
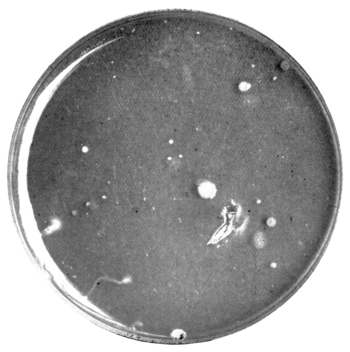
| 17. | Plate culture from fresh puer | " 97 |
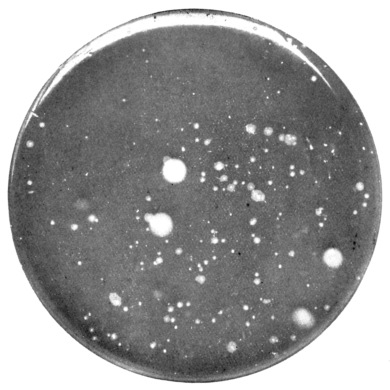
| 18. | " " puer wheel | " 103 |
| 19. | B. putrificus | " 103 |
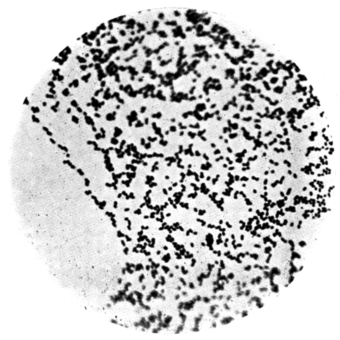
| 20. | Organisms in pigeon dung, × 1000 | |
| 21. | Pure culture of bacillus (d) from sweated skin | facing 106 |
| 22. | " " " (e) " " | " 106 |
| 23. | B. butyricus (Hueppe) | " 112 |
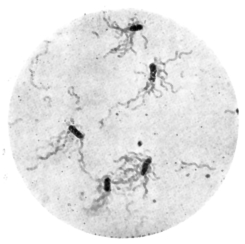
| 24. | Spirillum volutans (Kutscher), stained to show flagellæ | " 112 |
| 25. | " " " unstained preparation | " 112 |
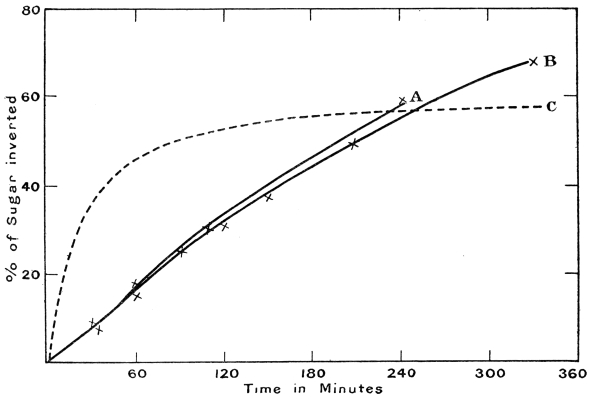
| 26. | Curves showing rate of hydrolysis | |
| 27. | Organisms in bran drench, × 1000 | |
| 28. | Chains of bran bacteria, × 1000 | |
| 29. | Bran fermentation, advanced stage | |
| 30. | B. furfuris, α | facing 269 |
| 31. | B. furfuris, β | " 269 |
| 32. | Cultures of α in glucose gelatin | |
| 33. | Gas curve (B. furfuris) |
The present volume is the outcome of a desire to preserve the numerous notes which I have made during over twenty years’ work at the practical and scientific study of bating. It has been my wish to complete the investigation of this important process in leather manufacture, for, as Lord Allerton has paradoxically remarked:1 “Good leather is made before the skins go into the tan liquor at all,” but owing to circumstances having drawn me more and more to the commercial side of the business, I have been compelled to abandon this project.
When learning the trade as an apprentice every fault in the leather was attributed to this part of the work, and the troubles and miseries of the “puer shop” first caused me to take up the study of puering. I was determined to know the causes underlying the process. Puering is not only a filthy and disgusting operation, but is prejudicial to health, and in the nature of it is attended by more worry and trouble than all the rest of the processes in leather making put together.
By giving a résumé of the work done up to the present time, I hope to be of assistance to the younger generation of Tanning Chemists, to some of whom is reserved the privilege of carrying on the work.
I think it may now be said, at any rate, that the solution of the problem of constructing an artificial bate on scientific principles, which will replace the present crude methods, is well within sight. The principal obstacles are, on the one hand, the inertia of English manufacturers; on the other hand, the class of labour employed in puering is not of the highest order of intelligence. Innovations in most things are resisted, partly because they necessitate changes in the method of working, and partly because of the innate conservatism of human nature. It is certainly a significant fact that although most of the pioneer work on this subject was done in England, the practical side has been taken up in Germany, and by freely spending money on large trials in the works they have enabled the manufacture of artificial bates to be developed on a commercial scale.
In 1886, while studying Chemistry under Professor Frank Clowes, I began to examine microscopically the various liquors of a light leather factory, and more especially the bran drenches. At that time I knew nothing of bacteriology, for the simple reason that little but pathological work in this line was being done in England. Through the kindness of Professor Clowes, I obtained an introduction to Mr. Adrian Brown (now professor at the University of Birmingham), and in his laboratory at Burton-on-Trent I saw the first pure cultivations of Bacterium Aceti which he had isolated, and of which he had completely studied the chemical action.2 I had there an opportunity of seeing the methods and apparatus employed. I also had the benefit of Mr. Brown’s advice in commencing a systematic study of the process of drenching. Professor Percy Frankland, then at Dundee, advised me further as to the microscope and other matters. My warmest thanks are due to these friends for directing me in the right way.
As a result of my first investigations, on December 11, 1889, I read a short paper entitled “Methods of Bacteriological Research—with some account of Bran Fermentation,” before the Society of Chemical Industry.
The way in which this paper was received led to a further research into the nature of bran fermentation in conjunction with Mr. W. H. Willcox, B.Sc. (now Senior Analyst to the Home Office), by which the action of the bran drench was thoroughly investigated, and the results published in the Journal of the Society of Chemical Industry, May 31, 1893.
This was followed, on June 30, 1897, by a paper “On a Pure Cultivation of a Bacillus Fermenting Bran Infusions,” also in conjunction with Dr. Willcox.
In 1898, in reply to a publication of Director Eitner, of Vienna, I published in the Leather Trades’ Review (November 15), a résumé of the whole subject, entitled “The Rationale of Drenching.”
Already, in the first paper above named (“Methods of Bacteriological Research”), I had called attention to the bacteria of the bate prepared from dogs’ dung, and in a paper entitled “Fermentation in the Leather Industry,”3 developed this aspect of the subject, and first pointed out the influence of enzymes in bating. I therefore decided to study the phenomena occurring in the bate in the same way as I had studied drenching.
The work was begun in 1895, and, as it was likely to occupy an indefinite time, the first instalment, entitled “Notes on the Constitution and Mode of Action of the Dung Bate in Leather Manufacture,” was published November 30, 1898; while “Further Notes on the Action of the Dung Bate” was published on November 30, 1899.
In these papers I indicated the lines on which a culture of bacteria might be practically applied to the bating of skins, and gave the composition of a liquid which, while acting as a nutrient medium for the bacteria, contained at the same time most of the active chemical compounds of dog dung.
Meanwhile, Dr. Popp and Dr. Becker, in Frankfort a/M, were investigating independently the bacteria of dog dung, and conceived the idea of employing them commercially. My dear friend Franz Kathreiner, of Worms,4 put me in communication with these gentlemen, and we were thus enabled to work in conjunction. As a result of our combined labours, an artificial bate, called “Erodin,” was put upon the market. This will be fully treated of in the chapter on Artificial Bates.
I shall give first a short account of bating, and then sum up as briefly as possible the present state of our knowledge of the process, afterwards giving an account of the more important of the various patents which have been taken out for artificial bates.
Although the book is divided into separate sections for convenience, it is obvious that we cannot separate chemistry from physics, nor bacteriology from chemistry, nor enzyme action from all three.
My own papers are printed as read. The Bibliography does not profess to be complete, but includes most of the works consulted.
No one realizes more than I how incomplete the work is, and how much research still remains to be done in order to complete it.
Thanks to the efforts of the Leather Industries Department of the University of Leeds, and the Technical College of the Leather Sellers’ Company, Bermondsey, the era of Rule of Thumb is passing, and there is little doubt that the work that is being done in these institutions will be translated into practical use in the factories by the coming generation.
I wish to express my special thanks to Mr. Douglas J. Law, and to Dr. H. J. S. Sand for assistance in preparing the notes for publication, to Dr. J. Gordon Parker, Director of the Leather Sellers’ Technical College, London, for the description of the bating of hides, and to Professor Kràl, of Prague, for some of the photographs of bacteria.
JOSEPH T. WOOD.
Nottingham : January 1912.
“Beizen sind Stoffe die mit dem Kalk nicht nur ein chemische Verbindung einzugehen im Stande sind, wodurch derselbe löslich und somit vollständig unschädlich wird, sondern auch den gegenseitigen festen Schluss der einzelnen Wandungen der Zellenelemente mechanisch lockern, den ganzen Bau der Haut in sich nachgiebiger machen und so die Verschiebbarkeit der einzelnen Hautgebilde erhöhen.”—J. C. H. Lietzmann, 1862.
“Bates are materials which are not only able to enter into chemical combinations with the lime, whereby it becomes soluble, and is thereby rendered completely harmless, but they also mechanically loosen the cementing substance of the separate cell elements, and render the whole structure of the hide more pliable, and thus increase the mobility of the various parts.”
The object of bating or puering is to render the skins, and the resulting leather, soft and supple. Skins which have undergone the liming process, must be thoroughly freed from lime before going into the tan liquors, and, for light and soft leathers, they must be reduced or “brought down,” so that the elasticity or resilience of the skin fibres is got rid of, and the skin, when tanned, can be stretched without springing back. This is usually done in the case of light leathers, by passing the skins through a bate or puer, composed of an infusion of dogs’ dung in water at a temperature of 35° to 40° C., until the required result is obtained. This condition is known to the workman by the feel of the skin. A good indication is, that the skin when “down” retains the impression of the thumb and finger if squeezed. A properly puered skin, when dropped on the floor, will also be perfectly flaccid, the folds lying closely together. It may, however, be said that it is only by experience and a kind of instinct that the exactly correct condition of the skin can be judged.
I know of no very early works on leather manufacture giving an account of bating. It was a “secret process,” and the results obtained depended almost entirely upon the judgment of the operator; and this judgment was frequently in error, owing to the fact that he did not understand what took place in the bate.
The earliest account I have been able to find is in a book5 in the possession of Mr. Seymour-Jones, of Wrexham, entitled “The Art of Tanning and Currying Leather, with an Account of all the Different Processes made use of in Europe and Asia for Dying Leather Red and Yellow, Collected and Published at the Expense of the Dublin Society, to which are added Mr. Philippo’s Method of Dying the Turkey Leather as approved of by the Society for the Encouragement of Arts, etc., and for which he had a Reward of £100, and their Gold Medal, for the Secret. Also the New Method of Tanning invented by the late David Macbride, M.D., London. Reprinted for J. Nourse, on the Strand, Bookseller to His Majesty, 1780.”
In the chapter entitled “Alumed Calf Skins for Bookbinding” (p. 138), after the limed skins have been fleshed, the writer continues—
“To alum them, put into a large vat, three or four pails of dogs’ turd (this dogs’ turd is called alum); on this they fling a large pail of water to dilute it; this done, the workman goes into the vat, and, with his wooden shoes, tramples it, filling the vat half full of water. The alumer, on his part, pours water out of his boiler into this vat, mixing it with the cold water, after which he flings in the skins, stirring them and turning them for some moments with great sticks.”
The work is described pretty much as now practised, the puer tub being kept at a uniform heat by constantly taking out liquor, heating it, and returning it. But neither these goods, nor morocco, are put through a “drench” after the puer, as we do now; they are scudded on the flesh, and well washed several times in clear water before being tanned.
Morocco Leather (p. 204).—The dry skins are soaked three or four days, “pared” on the beam, and unhaired in weak lime pits one month. At Nicosia they put the skins into lime, reduced to powder, for twenty days in summer, or twenty-five to thirty in winter; out of the lime, the skins are well washed and drained. The author continues—
“The river work finished, the skins are put into the dogs’ confit, or mastering; for every four dozens of skins they add one bucket of dogs’ excrement, containing fourteen or fifteen quarts, which is worked up with their hands into a kind of pap and well diluted. The skins are flung in, stirred and worked in the mastering for some minutes, then turned and left to rest.
“They remain about twelve hours in the mastering, which opens them, and takes off the rawness, disposes them to relax, fill and ferment. This excrement, by its alkaline parts, also cleanses them and takes out the grease, which would hinder them from taking the colour. I shall hereafter speak of bran mastering.
“At Diarbekir, they make use of these masterings in a different way. Whilst the skins are drying, they fill great hollows made in the earth, like our lime pits, with dogs’ dung, which is diluted to the consistence of honey, or of thin pap, in which they soak the skins for eight days in winter, and three in summer, treading them each day with the feet. They are taken out of this fecal matter, and well washed with fresh water, after which another mastering is made with bran diluted with water, in which the skins are soaked six days in winter, and three in summer, observing to tread them each day with the feet, the same as in the dogs’ mastering; they are then taken out, washed in fresh water, and prepared for dyeing.”
From these primitive methods the majority of tanners, even of the present day, have departed but little, except that, instead of stirring the goods and liquor with a stick, a paddle is used, and the process thus shortened considerably. Before describing the operation of puering, it will be best to describe the preliminary washing of the skins, because, although limed skins may be entered direct into the puer and brought down quite satisfactorily, in this case more puer and more time are required, hence it is usual to get rid of the bulk of the lime by washing them in water before putting them into the puer. In some cases very dilute hydrochloric or other acid is used, in order to shorten the time of watering.
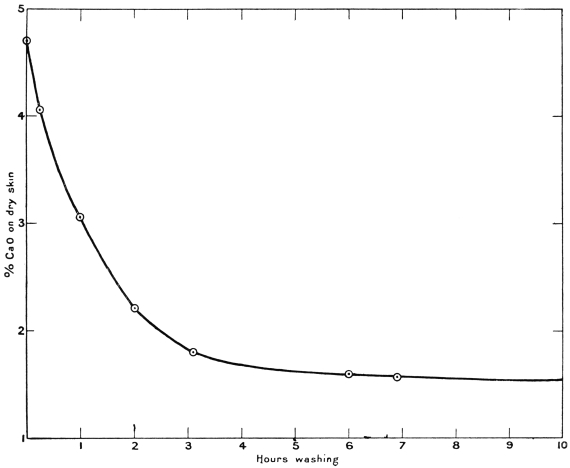
It is now well known that lime cannot be entirely removed from skins by washing in water, no matter how long the washing be continued. A limed skin containing 4·6 per cent. CaO, calculated on the dry weight, was found to contain after washing—
Time of washing | Per cent. CaO on dry skin | Per cent. lime removed | |
1 | hour | 3·05 | 34 |
2 | hours | 2·20 | 52 |
3 | " | 1·75 | 62 |
6 | " | 1·55 | 66 |
7 | " | 1·55 | 66 |
24 | " | 1·50 | 67 |
If the washing be continued, a less amount of lime is removed in each successive period of time, so that it is evident a point is soon reached at which it becomes a waste of time to continue the washing. In practice this point is reached in about two hours. The progress of the washing is best shown by the curve in the diagram (Fig. 1), in which the ordinates represent the percentage of lime (CaO) in the dry skin, and the abscissæ time in hours. It will be seen that the character of the curve is a hyperbola; such a curve only approaches a line (representing in the example chosen about 1 12 per cent. of lime) asymptotically—in other words, it is impossible to wash out all the lime except by an infinite number of changes of water, since each washing removes a less amount than the previous one. The above is a typical case of the washing of limed grains from the splitting machine in the manufacture of skivers. These contain from 4 to 5 per cent. of lime on the dry skin, and, after washing in water in a paddle for six hours, analysis shows them to contain still about from 1·5 per cent. to 1·9 per cent. of lime (CaO).6
The effect of washing depends on the character of the water (hard or soft), and also on its temperature. Hard waters should have a small quantity of clean lime added to them before entering the goods, in order to remove dissolved CO2, which, by carbonating the lime on the surface of the skins, renders the grain harsh and the subsequent puering difficult.
With regard to temperature, it is preferable to employ cold water until the bulk of the lime is out, since this dissolves more lime than warm water. 100 c.c. of saturated lime water, at 10° C., contains 0·134 grm. CaO. 100 c.c. at 40° C. contains 0·1119 grm. CaO. Moreover, a comparatively small rise of temperature causes a considerable decomposition in a fully limed skin, by which the skin substance is rendered more soluble, and consequently lost for the purposes of the tanner. For a fully limed skin the limit of temperature is about 82° F. (28° C.), whereas a skin free from lime or alkali may be submitted to a temperature of 120° F. (49° C.) without damage to the fibre. Lamb prefers a short washing of half an hour in water at 35° to 38° C., for the reason that the increased temperature causes the goods to become more flaccid, but this condition is attained at the expense of the loss of skin substance we have mentioned.
In order to save time and water, the following method is adopted. The goods are measured by means of a cubical truck on wheels (Fig. 2); it holds 250 kilos of wet skins (550 lb.). Four of such trucks are placed in a wash wheel, and a stream of water from a 1-in. pipe turned on. The goods are run from three-quarters of an hour to one hour; the water is then stopped, and 4000 c.c. of commercial hydrochloric acid (18° Bé.) is run in slowly, in a very diluted state, through a perforated lead pipe. After all the acid has run in, the wheel is run for half an hour, then water turned on again for half an hour in order to wash away the calcium chloride produced. During this last washing, hot water is admitted at the back of the wheel through a perforated pipe, in order to raise the temperature of the goods, so that they do not enter the puer wheel in a cold condition, and thus lower the temperature of the puer liquor. The goods are now ready for puering.
The chemical action of the acid is a very simple one, and is expressed by the formula—
It will be noted that the quantity of acid used is only about one-tenth of that required to neutralize completely the lime in the skin. If a greater amount of acid be used the skins begin to absorb acid before the lime in the interior is neutralized; but, provided the lime on the surface of the skins is removed, that remaining in the interior is forced out during the course of the bating process (see Chapter III.). If more acid were used, then it would require to be added so slowly that time would be wasted unnecessarily in doing the work.
With regard to the use of other acids for deliming, Lamb considers formic acid preferable to hydrochloric, and states that the removal of lime is more effectively accomplished by its use. He attributes a pulling down action to the calcium formate produced, so that less formic acid is required than would be equivalent to hydrochloric, merely regarded as a solvent for lime. We shall refer to this point again in Chapter II. It may be stated here that the author has obtained better results by the use of a mixture of formic and acetic acid, in equal proportions, than from either acid alone, but that the cost of working is considerably greater than with hydrochloric acid.
Lactic acid is in fairly common use as a deliming agent. In practice 1 per cent. lactic acid (50 per cent. strength) calculated on the wet weight of pelt, is used, added gradually in small quantities. This does not remove the whole of the lime, but sufficient for the skins to puer quickly.
Some tanners measure their acid per dozen skins. In a case which came under my notice, 1200 c.c. 90 per cent. formic acid were being used for ten dozens medium goat skins at a temperature of 35° C., time 1 14 hours.
The following table, due to Professor H. R. Procter,8 gives the cost of dissolving 1 lb. of lime at present approximate prices of the various acids. The dissociation constant K shows their relative “strength;”9 the equivalent, Eq., the weight in lb. of 100 per cent. acid required to dissolve 28 lb. of lime. Ordinary wet limed hides, unhaired and fleshed, contain only about 4 lb. lime per 1000 lb. wet weight.
Cost of Acids to remove 1 lb. Lime.
Acid | Eq. | K | Cost per cwt. | Strength per cent. | Cost for 1 lb. CaO | |
s. | d. | d. | ||||
| Hydrochloric | 36·5 | say 200 | 3 | 3 | 31·5 | 1·4 |
| Sulphuric | 49·0 | " | 4 | 0 | 95·0 | 0·8 |
| Oxalic | 63·0 | 0·1 | 30 | 4 | 99·0 | 8·1 |
| Formic | 46·0 | 0·0214 | 35 | 0 | 87·4 | 7·0 |
| Lactic | 90·0 | 0·0138 | 26 | 0 | 49·7 | 18·0 |
| Acetic | 60·0 | 0·0018 | 18 | 0 | 40·0 | 10·0 |
| Butyric | 88·0 | 0·00115 | 21 | 0 | 82·8 | 8·0 |
| Boracic | 62·0 | 0·00000001 | 27 | 0 | 99·0 | 6·5 |
It will be seen from the above table that sulphuric is the cheapest acid to use; but, owing to the insoluble nature of the calcium sulphate, it is best to use hydrochloric acid. Procter has suggested using a mixture of sulphuric acid and common salt in molecular proportions, so as to avoid the iron which is generally present as an impurity in commercial hydrochloric acid. The author has tried this, and found it perfectly satisfactory. For a complete discussion of chemical deliming, see Procter’s “Principles of Leather Manufacture,” chapter xiii.
Quite recently butyric acid, which is now being manufactured on a commercial scale by the process of Dr. Effront (Brussels), has come into use for deliming.10 and is likely to become a useful acid, as it can be manufactured cheaply. According to Parker, it dissolves less skin substance than formic, acetic or lactic acids, which observation is confirmed by Thuau. The quantity required per 100 lb. of ordinary washed sheep skin is about 12 lb. of the 80 per cent. acid. It is used in exactly the same way as other acids, i.e. it should be added in small quantities, suitably diluted, at successive intervals of time.
Acid salts are also used for deliming, and Procter suggested sodium bisulphate NaHSO4 (“Principles,” p. 155). More recently, sodium bisulphite (NaHSO3) has been recommended;11 the skins are tumbled for half an hour with a dilute solution, which combines with the lime according to the following equation—
if sulphides are present the reaction is—
After this treatment the theoretical quantity of hydrochloric acid, diluted with plenty of water, is run in through the axle of the tumbler, and the skins run another half hour. The reactions are—
(1) CaSO3 + 2HCl = CaCl2 + SO2 + H2O
(2) Na2SO3 + 2HCl = 2NaCl + SO2 + H2O
The sulphurous acid evolved in this way slightly swells the skins, and also removes stains due to the alkaline polysulphides; it is also said to remove salt stains.
| CH2 CH2 | O | |
| CH2 CO |
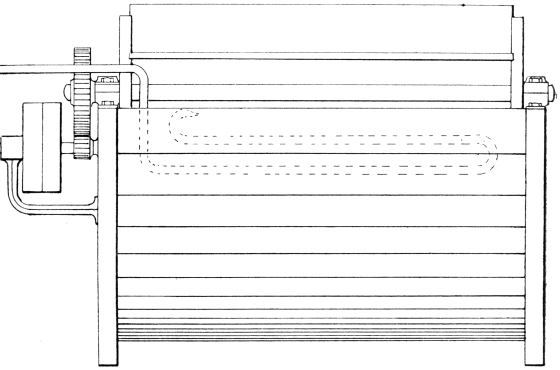
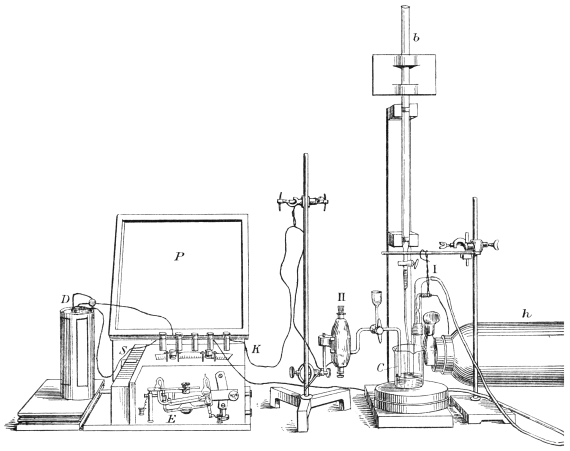
After washing and deliming, the goods, warmed to a temperature of 90° to 100° F., are now measured in the truck above described, and put as quickly as possible into the bating paddle. This is constructed to carry two trucks. The figures (Figs. 3, 4 and 5) show a modern paddle adapted for puering light skins, such as goat, sheep, splits (grains), either with dung or with an artificial bate.
The inside dimensions are: length, 4 ft. 6 in.; width, 4 ft.; depth, 2 ft. 6 in. A copper pipe of two coils is provided at the back, through which steam is passed for maintaining the temperature of the wheel—the goods are prevented from coming in contact with this pipe by a perforated board. Another steam pipe, jointed so that it may be turned in and out of the paddle, is provided for heating up the wheel to the required temperature before beginning the bating. This pipe is not shown in the drawing. The thermometer can be read while the paddle is in motion. A suitable speed to keep the goods in motion is twenty revolutions per minute. The whole wheel is covered in, for the purpose of preserving the heat, and also to exclude light and air, which are prejudicial to the action of the bacteria. Such a paddle, or wheel, has a capacity of 200 gallons liquor, or 900 litres, and will bate 25 dozens of average grains, weighing in the wet state 1100 lb. or 500 kilos.
The dog dung, which is usually obtained from hunting kennels, and of which the composition is given in Chapter II., is simply diluted with water in the paddle;13 five buckets, weighing about 165 lb., are required for the above quantity of goods.14
The amount used depends on the state of the goods, the prevailing weather, and, further, on the quality of the puer.15 When the temperature is maintained at 40° C., ordinary limed goods, which have been washed, come down in from one to three hours.16
During puering, the lime soaps in the skin are decomposed, and the fat set free—in an estimation of the fat, 0·151 per cent. on the wet skin was found before puering, and 2·48 per cent. after puering. The fat thus set free can be removed to a great extent by scudding; but in modern practice this is not pushed to extremes, as the grain is liable to be damaged by excessive scudding. The fat is afterwards removed from the dry leather by means of benzine or other solvent, employed in a suitable apparatus.
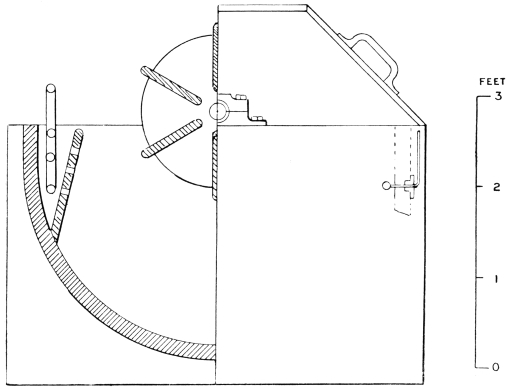
Scudding.—When the goods are “down,” they are ready for scudding. This operation in the case of grains is performed either by hand over the beam, or by means of a special scudding machine.
The scudding machine figured (Fig. 6) was invented by Sir John Turney in 1880,17 and is the only type of machine which will safely scud split skins. It will readily be understood that the usual type of scudding machine, in which a spiral knife is used, would tear the delicate structure of the grain. The Turney scudding machine works with hard, circular brushes, which revolve against a wooden roller carrying the skin. At the same time a continuous stream of water, at a temperature of 110° F., is sprayed upon the skins from a perforated pipe.
After scudding, the skins are washed in cold water for about ten minutes. This checks the action of the puer, and also washes away any loose dirt. They are then ready to pass on to the drench (Chapter IX.), or to the pickling process, or direct to the tan liquor, according to the kind of leather required.
The Bating of Hides for Harness or Dressing Purposes.—The hides intended for harness leather, or for dressing purposes, are usually bated with hen or pigeon manure. The exact process varies in different tanyards, being dependent upon the condition of the pelt prior to bating, as to whether it is bated directly after fleshing or receives a preliminary deliming. The process is also dependent upon the amount of bating action required for the specific purpose prior to tanning, but the following three methods are in actual use, and may be taken as typical methods:—
Harness Backs.—About a hundredweight of hen manure is put into a suitable tub or vat capable of holding about fifty gallons; some thirty gallons of water, at a temperature of about 100° F., is poured on to the manure, and the whole is thoroughly stirred up by means of a wooden stick or plunger, working the manure about in order to mix it to a consistence of thin soup. The vat is now stored in a warm place, so that the temperature is kept about 70° F.—it is customary to store this in the boiler-house, or some similar warm room. It should be stirred two or three times a day, until it begins to show signs of working; two days is generally sufficient for this.
A pit is now prepared, with the necessary amount of water; and, by the aid of a steam pipe, or other similar means, the temperature of the water is raised to 70°, and the contents of the bate tub are now added. This should be poured into the pit through a sack, or a canvas filter, so as to keep back stones, solids, feathers, and other extraneous matters. The pit is now thoroughly plunged, and the goods entered; these should be handled up three to four times a day for two days. At the end of two days the goods are usually scudded, by working the back on an ordinary unhairing beam with an unhairing knife. They then go into a fresh pit containing a new solution of bate made up in the same manner, and, when sufficiently bated, which generally takes about three days, the goods are removed from the pit, scudded again by working them over with a slate scudding tool, when they are ready for tanning. Some tanners give them a bath of boracic acid prior to their going into the tanning liquor; this has the effect of materially brightening the colour. The second pack of harness backs go into the same liquor—which must be re-warmed to a temperature of 70°—in which they are handled as before for about two days, and are finished off in a new liquor made as above described.
It will be understood, therefore, that in a tanyard, working regularly, the pack of backs go first into an old bate for two days, and are then finished off in a freshly made one; so that each pack gets two days in an old pit, and one or more days in the new. The bate in the tub may also have a second water, or be strengthened.
Dressing Hides.—For these goods a somewhat stronger bating effect is required, and pigeon manure is therefore frequently used. The preparation of the bating liquor is as described above, both as regards quantity and temperature of the water and time of maceration.
The whole process may be carried out exactly as described for harness backs, but the following is an illustration of another method in practical use. Imagine four ordinary bating pits, worked in a round. The first pit is very old, having had three packs of hides through it; the second having had two packs, and the third one pack; and the fourth is made new. The goods, after unhairing and fleshing, and washing, are put into the first pit quite cold, are handled in this three times on the first day, and afterwards put into the second pit, the temperature of the second pit having been previously raised to 65°—the cold pelt going into this pit reduces the temperature to about 60°. They are handled in this pit twice, and remain overnight; and the third day they go into the third pit, the temperature of which is raised to 70° before the goods go in. They are raised, scudded thoroughly on the grain, and new liquor is made from new bating liquor prepared as described above; the goods are now entered, handled at least twice a day, and, if insufficiently bated, are left overnight. By the next day they will probably be sufficiently “down” to permit of them going on to the scudding beam and thence on to tanning.
In the event of any hide being insufficiently bated—in other words, if the “bone” has not completely disappeared in the inner part of the hide—they are returned once more for three or four hours, the temperature of the pit being raised to 70° with a steam coil.
By this system, which is known as a system of “sets,” it will be noticed that the pits are worked in a round of four; the first pit, or oldest, being used more or less as a wash pit, the actual bating taking place in the second, third and fourth pits. No pit has more than four packs passing through it, and the temperature never rises above 70°, or, at the outside, 75°. The reason of the comparatively low temperature is due to the fact that in pit bating, if the temperature is raised above 75°, the grain of the hide becomes affected before the internal “bone” is sufficiently soft, which results in a tender grain, which considerably affects the resulting staining or dyeing which the leather usually receives before it is put on the market.
The Third Process.—This process is suitable for either harness or dressing hides, and differs from the above-described process only in the fact that instead of bating the hides in pits, a latticed drum is used in order to keep the hides in constant motion. This consists of a large pit, in which a latticed drum revolves two-thirds immersed in the liquor. The bate liquor is made up as usual, and the temperature of the liquor raised to 75° or 80° F.; the hides are then placed in the latticed drum, which revolves at from four to six revolutions per minute, and, if the goods are put in the drum in the morning, and if the temperature is kept at about 70° F., the hides are generally sufficiently bated by evening. They may then be taken out of the drum, scudded, and left suspended in boracic acid overnight, and can then be taken to the liquor next morning.
Where there is considerable motion, even the heaviest hides will stand a temperature of 75°, and even 80°. Some tanners who use the latticed drum method, do not place their hides in the drum until the afternoon; they are drummed for about an hour in the cold bate, and left overnight; next morning the temperature of the bate is raised to 75°, and the drum started; they are then removed from the drum towards the afternoon, when sufficiently “down” to admit of them passing forward into the next process.
The above are three typical methods in common use in England, but many modifications of these processes exist. Everything depends upon the amount of lime in the hide, on the number of haulings or handlings given to the goods, and the temperature or the conditions under which the hides are bated, so that no hard-and-fast lines can be laid down. Much also depends upon the method of liming used, prior to the bating. It must be borne in mind that the bating of hides, and even of kips, differs materially from the bating of goat and sheep skins, as one has to deal with double or even treble the substance. If the bate is worked at a temperature of even 70° or 75°, there is the danger of the grain being seriously affected before the bate has penetrated to the interior of the pelt, unless the goods are kept in constant motion.
Instead of a latticed drum, large paddles are used in some factories. If the pit is large enough the paddle causes both hides and liquor to revolve, and thus gives the required movement necessary for a regular and even “bating effect” to be obtained.
“And now, as we cannot understand the frame of a Watch, without taking it into pieces; so neither can Nature be well known, without a resolution of it into its beginnings, which certainly may be best of all done by Chymical Methods”—Jos. Glanvill, 1668.
Professor H. R. Procter, in his “Principles of Leather Manufacture,” 1903, p. 153, has given a very complete account of chemical deliming, and also of the bating and puering processes. Meunier and Vaney, “La Tannerie,” 1903, give a general review of our knowledge of these processes up to that date. These accounts are extremely useful, but treat the subject in a general manner.
I propose to treat of the particular case of puering by means of an infusion of dogs’ dung in water, as I believe this will give an insight into all the other bating processes.
So far as the purely chemical action of the bate goes, it consists in the solution of the lime contained in the skins,18 thus setting free a certain portion of skin substance which was combined with the lime, and the subsequent solution of more or less of this skin substance.
If a fresh puer be made and boiled for half an hour, then allowed to cool to 95° F., it will be found to remove the lime from the skin in a very similar way to an ordinary bate, but it has not so rapid a reducing action as an unboiled bate. In this case both bacteria and enzymes are destroyed, so that the action may be put down to the chemical constituents of the bate.
Chemical Composition of the Puer.—The mineral constituents of the fæces, both in dogs and human beings, are well known, owing to the study of the processes of digestion and nutrition in physiological laboratories; but the organic constituents are yet little known, and the sum of the weight of those at present estimated is far from the total of these matters present.
It is a mistake to suppose that the fæces represent the residue only of the food taken. Strassburger19 estimates that bacteria alone account for nearly one-third of the dry matter. In addition, the intestinal mucous membrane is an important excretory channel for lime, magnesia, iron and phosphoric acid, as has been shown by analyses of fæces from men and dogs, during prolonged fasting.20 Even when no nitrogenous food is eaten, the dry fæces always contain from 4 to 8 per cent. of nitrogen; in a dog fed on meat it amounts to 6·5 per cent.
In making a chemical analysis of the fæces, the mineral matter is estimated in the ash in the usual way for ash of organic substances, with special precaution, on account of the phosphates present. The material is first charred, the salts removed by acetic acid; the acetic acid solution is then decanted, the residue washed with distilled water, and the combustion then completed. The acetic solution and washings are added to the final ash, the whole evaporated to dryness, and gently ignited to decompose the acetates. (Cf. Bull. 46, U.S. Dept. of Agriculture, Washington, 1899.)
In estimating the fats, the dry puer is ground up with sand, then extracted with ether in a Soxhlet. The fats are present in four states:—1, neutral fats; 2, free fatty acids; 3, alkali soaps, soluble in ether; 4, small quantities of lime and magnesia soaps remaining in the residue in the Soxhlet. For details as to the separation and estimation of these, Lewkowitsch’s “Oils, Fats and Waxes” (Macmillan) should be consulted.
The separation and estimation of the organic constituents in the puer, is one of the most difficult problems of physiological chemistry, and to include the methods used would demand a special treatise beyond the scope of the present volume. Hoppe-Seyler’s “Physiological Chemistry,” and Allen’s “Commercial Organic Analysis,” vol. iv., may be consulted; and there is a mass of useful information in Dr. René Gaultiers’ “Précis de Coprologie Clinique” (Paris, Baillière et Fils, 1907).
It is proposed to give here the results of a number of analyses of dog dung made by the author and others, and to discuss the action of the various bodies upon the skins. Many more experiments and much research requires to be done, before the complete action of the bate is clear, but only by the method of experiment with the various bodies present can the problem eventually be solved.
Analysis of raw puer from hunting kennels (Quorn) showed the approximate composition of 1000 grm. of raw puer, containing 150 grm.21 of dry matter to be as follows:—
| Grammes | ||
| Sodium chloride and sodium sulphate | 2 | ·1 |
| Sodium ammonium phosphate | 14 | ·0 |
| Earthy phosphates, principally Ca3(PO4)2 | 33 | ·6 |
| Ferric phosphate | 0 | ·87 |
| Calcium sulphate | 1 | ·94 |
| Silicic acid | 3 | ·40 |
| Calcium in solution as CaO | 1 | ·42 |
| Total P2O5 in solution | 4 | ·00 |
| Non-volatile acids (as lactic) | 3 | ·00 |
| Volatile acids (as acetic) | 2 | ·20 |
| Amines (as ammonia) | 6 | ·20 |
| Enzymes | 3 | ·66 |
| Nitrogenous bodies not included in the above, consisting of complex amido-acids, leucin, tyrosin, xanthin, and other purine bases, indol and skatol | 17 | ·00 |
| Organic matter, cellulose, etc. mostly insoluble, including fats and soaps | 70 | ·00 |
On the dry matter the amounts extracted by various solvents were:—
Per cent. | ||
| Water | 24 | ·03 |
| Alcohol | 22 | ·27 |
| CS2 | 15 | ·57 |
| C2HCl3 (Tri-chlorethylene) | 14 | ·45 |
| Petrol. ether | 13 | ·23 |
No carbohydrates (sugars) capable of reducing Fehling’s solution were found in the puers examined.
The ash22 of the dry fæces from dogs fed on a flesh diet varies from 20 to 34·27 per cent., with the following percentage composition:—
| Silica | 7 | ·04 |
| CO2 | 4 | ·62 |
| SO3 | 7 | ·37 |
| Fe2O3 | 4 | ·22 |
| CaO | 25 | ·29 |
| P2O5 | 26 | ·41 |
| MgO | 15 | ·52 |
| Cl | 1 | ·50 |
| Alkali | 5 | ·53 |
In a dog weighing 30 kilos., fed on a mixed diet of 500 grm. meat and 200 grm. starch, 78·6 grm. dry fæces were obtained containing 23·76 per cent. ash of the following percentage composition:—
| CaO | 22 | ·3 |
| P2O5 | 25 | ·4 |
| Fe2O3 | 10 | ·6 |
| MgO | 9 | ·8 |
| SO3 | 5 | ·0 |
| Cl | 0 | ·2 |
| Alkali | 1 | ·1 |
| Insol. in HCl | 21 | ·8 |
It is obvious that the composition of the fæces will vary with the food given, and in this connexion it may be stated that the puer used was obtained from dogs fed on a mixture of boiled horseflesh and oatmeal porridge, about equal quantities. Sometimes charcoal and cabbage are mixed with the food, in this case the puer is very dark coloured. The dark colour so produced is to be carefully distinguished from the dark colour caused by decomposition of the puer.
We cannot leave out of account the urinary products, which, in the case of dung from hunting kennels, are always present, though in varying quantities.
The total nitrogen in 1520 c.c. of urine was found23 to be 15·9 grm.
Each 100 grm. total nitrogen was distributed as follows:—
| Urea | 85 | ·9 |
| NH3 | 4 | ·1 |
| Creatinine | 3 | ·3 |
| Uric acid | 0 | ·5 |
| N in other compounds | 5 | ·7 |
The inorganic compounds were:—
Per cent. | ||
| Sulphur (as sulphate) | 3 | ·31 |
| P2O5 | 3 | ·98 |
| Cl | 6 | ·30 |
The urine of carnivora is acid, that of herbivora is neutral or alkaline.
Urea, which is the chief nitrogenous constituent, is very quickly fermented by several species of bacteria, and entirely decomposed into ammonium carbonate, so that it is this latter body which acts in the bate.
| CH2 | NHC, H5O | ||
| COOH |
The analysis of the lion’s dung gave the following results:—
Per cent. | ||
| Water | 59 | ·2 |
| Ash | 21 | ·1 |
| Lime (CaO) | 10 | ·3 |
| Phosphates (P2O5) | 10 | ·67 |
| Organic matter | 19 | ·7 |
Dog dung contains some non-volatile bases of the purine group, such as xanthin (C5H4N4O2) and guanin (C5H5N5O), but the action of these bodies in the bate, has not been studied. It is also extremely probable that these and similar bodies are formed during the bating by a process of auto-digestion of various proteids.24 The quantity of fat in dry puer, amounts to 10–11 per cent. Cholesterol was present.25 Part of the fat is in the form of lime and magnesia soaps, but part in the form of an emulsion, which probably plays an important part in the puering process. The function of the fatty constituents of puer is one of the problems on which further research is necessary.
The colouring matters of the puer are nearly all derived from bile products. Most of them are found in the petroleum extract, which contains far more of the colouring matters than the chloroform extract.
The following bile colouring matters have been prepared by Merck, of Darmstadt: bilihumin, biliprasin, bilirubin, bilifuscin, biliverdin.26 So far as has been ascertained by the author, their effect in the bating is nil, but, in so far as they impart colour to the pelt, they are detrimental.
Experiments on the action of bile are given in the paper reprinted in Chapter VI.
Recently Eberle27 has proposed to add a proportion of gall juice (bile) to a bate containing pancreatic enzymes, with a view of energizing the latter. This part of the subject will be dealt with in Chapter V.
The Reactions of the Puer.—The organic acids are present principally in the form of the sodium or calcium salts. The intestinal juices contain sodium carbonate, but this is neutralized by lactic acid found in the intestines, so that the excreta usually have an acid reaction. The reaction of a fresh puer liquor is faintly acid to litmus, but this acid is immediately neutralized by the introduction of the skins, so that the rest of the bating process takes place in a neutral or alkaline medium, and the quantity of lime removed by the free acids is probably small.
The acidity of a filtered puer liquor made up freshly and titrated with N/5 soda, and phenolphthalein as indicator, was found to equal 10 c.c. N/1 acid per 1000 c.c. of bate. In the volume of liquor used, namely, 900 litres, this would neutralize only 250 grm. of lime (CaO).
The determination of the exact acidity or alkalinity of puer liquors by ordinary methods with indicators is not easy, as the end reactions are not sharp. The liquor above described was alkaline to methylorange and litmus, although acid to phenolphthalein, and the results also vary somewhat, according to the quantity of bate used for titration, and according to the strength of the acid or alkali used. The method used, both at the Trent Bridge laboratory and Messrs. Doerr and Reinhart’s laboratory at Worms, is as follows:—
The puer liquor is filtered through a Schleicher and Schulls hard filter, No. 602, 18 12 cm. diam.; 50 c.c. are taken, 4 drops of phenolphthalein solution (10 gr. dissolved in 300 c.c. alcohol) added, and titrated with N/5 acid or alkali as the case may be.
If the acidity be determined by adding excess of alkali and titrating back, a larger amount of acid is found than by direct titration. In the above-mentioned case, an indirect titration showed an acidity equal to 18 c.c. N/10 acid per 100 c.c. of liquor.
On adding the alkali to a fresh filtered puer liquor, a flocculent precipitate is produced, which appears to be due to the decomposition of proteid compounds of weak organic acids,28 the alkali uniting with them and setting the proteids free. As these compounds are undoubtedly decomposed by the lime in the skins, it seems probable that the acidity available for neutralizing lime may be greater than that shown by direct titration.
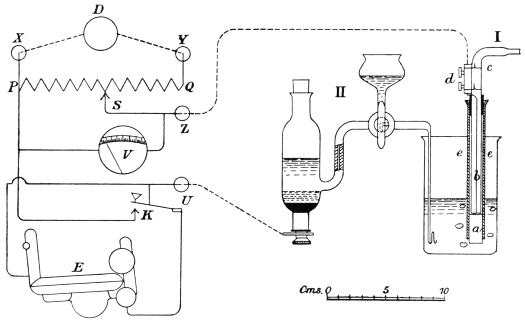
A method which was suggested by me for the estimation of the acidity of tan liquors may be applied also to the examination of the bate liquors, namely, the method by which the electric potential between a hydrogen electrode dipping into the bate liquor, and a standard calomel electrode, is used to determine the neutral point (for details see Chapter III.), using the potentiometer devised by Dr. H. J. S. Sand.29
Puer liquors, titrated in this manner with N/10 soda or HCl until a potential of 0·69 volts is reached (at which point phenolphthalein turns from colourless to red), gave the following results per 100 c.c. filtered liquor.
| No. | Description of Liquor | Reaction | c.c. N/10 | Alkali | Required for Neutralization | ||
| or | |||||||
| Acid | |||||||
1 | New puer before goods | Acid | 7 | ·4 alkali | |||
2 | The same after goods | Alkaline | 0 | ·57 acid | |||
3 | New puer before goods | Acid | 8 | ·1 alkali | |||
4 | The same after goods | Alkaline | 3 | ·25 acid | |||
5 | Used puer after goods | Alkaline | 5 | ·00 acid | |||
6 | Spent puer | Alkaline | 6 | ·6 acid | |||
Determinations made in this way are more accurate than the colorimetric method above described. Puer liquors after use are found by the electrometric method generally on the alkaline side, whereas such liquors show neutral to phenolphthalein owing to some action of the bate constituents on the indicator.
Puer Liquors in Electrometric Apparatus.
No. | P.D. Before Skins volt | c.c. of N/10 KOH required for 50 c.c. liquor to 0·69 volt | P.D. After Skins volt | c.c. of N/10 HCl required for 50 c.c. liquor to 0·69 volt | Description of Liquor | |||||
A1 | 0 | ·622 | 1 | ·95 | .. | .. | New puer | |||
A2 | .. | .. | 0 | ·747 | 1 | ·9 | A1 after 2 packs skins | |||
B1 | ·607 | 3 | ·7 | .. | .. | New puer | ||||
B2 | .. | .. | ·770 | 3 | ·1 | B1 after 3 packs skins | ||||
C1 | ·580 | 4 | ·3 | .. | .. | New puer | ||||
C2 | .. | .. | ·762 | 2 | ·8 | C1 after 2 packs skins | ||||
D1 | ·580 | 5 | ·2 | .. | .. | New puer | ||||
D2 | .. | 0 | ·8 | ·680 | .. | D1 after 1 pack skins (still on the acid side of 0·69) | ||||
E1 | ·570 | 6 | ·2 | .. | .. | D2 + 4 buckets more puer | ||||
E2 | .. | .. | ·720 | 0 | ·8 | E1 after 1 pack skins | ||||
F1 | ·610 | 3 | ·0 | .. | .. | New puer | ||||
F2 | .. | 0 | ·5 | ·680 | .. | F1 after 1 pack skins (still on the acid side of 0·69) | ||||
The potential at the neutral point, using an auxiliary electrode filled with N/1 potassium chloride, was 0·69 volts. Potentials below this point, therefore, indicate an acid reaction, and, conversely, potentials above 0·69 volts indicate an alkaline reaction. The apparatus is thus very useful in following the course of the bating process. The initial potential also enables the hydrion concentration of the bate liquor to be determined directly (see Chapter III.). It was found that during the bating of a pack of skins the hydrion concentration of the liquor diminished from 10-5·2 to 10-7·4 normal.
Such acidities as are found by this method, e.g. 7–8 c.c. N/10 acid per 100 c.c., are too great if made up by taking a solution of the free acids. The ionization must be almost entirely repressed by the presence of sufficient neutral salts of the same acids in the bating liquid, in order to obtain a similar result to the puer.
The table opposite gives the values found for a series of puer liquors, made with dog manure from hunting-kennels.
The reaction of puer liquors (expressed in c.c. of N/1 alkali or acid per 1000 c.c. of bate), in a series in which the washed goods were run for one hour in old puer, out of which goods had been taken, and then transferred to a freshly-made puer, was as follows:—
Bate | Before Goods | After Goods |
| Old puer | Neutral | 28 c.c. acid |
| Fresh | 10 c.c. alkali | Neutral |
| Old puer | Neutral | 40 c.c. acid |
| Fresh | 11 c.c. alkali | Neutral |
These figures were obtained by the ordinary colorimetric method described on p. 32.
Adding together the alkalinity of the bate run away, and the acidity of the new bate which is neutralized, the total for 1 and 2 = 38 c.c. N/1 acid, and for 3 and 4, 51 c.c. N/1 acid.
Multiplying by 810 (volume of bate less volume of dry skin), we find that the skins have neutralized 30·78 and 41·31 litres N/1 acid respectively. Since one litre of N/1 acid will neutralize 28 grm. of lime, it follows that the lime removed by the bate was in one case 862 grm., in the other case 1157 grm.
The 500 kilos, of wet washed grains introduced into the paddle contain approximately 0·5 per cent. of lime CaO, equivalent to 2500 grm. altogether. As will be seen from the figures given in table, p. 35, the free acids present in the fresh bate are only capable of neutralizing 10 × 810 c.c. of normal alkali, and this is equivalent to 225 grm. of CaO, or 9 per cent. of the total lime.30
In addition to this, a considerable amount of lime is removed from the skins which is not found afterwards in solution, but which is precipitated in an insoluble form during the course of the bating process. In an experiment to determine the amount of this insoluble lime, the following figures were obtained:—
Lime in Puer Liquor, Grams per Litre CaO.
| Before Skins | After Skins | |
| Soluble lime | 0·19 | 0·485 |
| Insoluble lime | 0·13 | 0·485 |
| ─── | ──── | |
| 0·32 | 0·970 |
It will be noted that the amount of insoluble lime has increased to a greater extent than the soluble lime, showing that part of the lime from the skins has been precipitated in an insoluble form. The increase of lime in the solution is 0·295 grm. per litre of soluble lime, and 0·355 grm. per litre of insoluble lime, a total increase of 0·650 grm.; that is, the ratio of soluble lime to insoluble lime in the original puer is 1·46 : 1. In the used puer it is 1 : 1, so that in the above case more than half the lime removed from the skins (54·5 per cent.) has been precipitated in an insoluble form, some of it in the form of calcium phosphate, and the remainder probably in the form of calcium oxalate. Crystals of calcium oxalate may be seen under the microscope in the puer residues.
The oxalic acid is produced by bacterial action, as has been shown by Zopf31 and Banning32; but it is difficult to estimate it in the bate, and a reliable analytical method of doing so requires devising.
A further 30 per cent. to 40 per cent. of the lime in the skins is removed by the chemical action of the complex amines of the organic acids, and by the mechanical effect of the paddle or drum, so that the final result is more or less as follows:—
Per cent. | |
| Lime neutralized by free acids | 9 |
| Lime dissolved by complex amines | 25 |
| Lime precipitated | 30 |
| Lime remaining in the skins | 36 |
── | |
100 |
The lime remaining in the skins is not in the caustic state, but principally in the form of neutral salts (see footnote to p. 24).
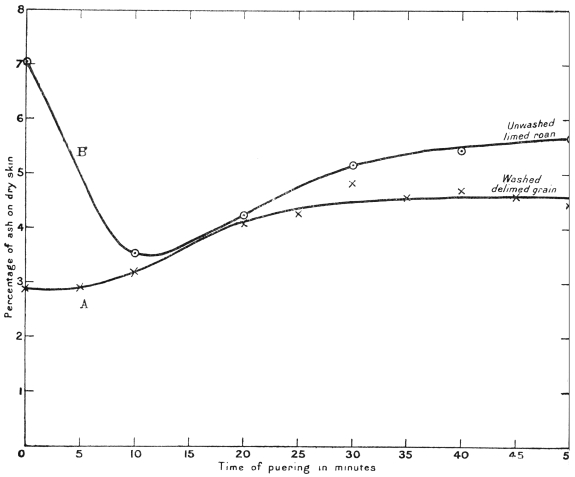
Some of these neutral salts appear to be absorbed by the skin during the course of the bating, for, on examining pieces of the same skin at frequent intervals during the operation, and determining the ash, it was found that a minimum point for ash content was reached in about ten minutes, after which the ash actually increased. The results are shown in the following table, and also graphically by the curves, Fig. 7—in which A is a “grain” previously washed as free from lime as possible; B, an unwashed “roan.” The effect of the absorption of inorganic matter is very noticeable in both cases. A part of the effect only is apparent since a certain amount of skin substance passed into solution.
Variation of Ash Percentage during Puering.
— | Ash, per cent. on Dried Skin | |
Time of Puering in Minutes | Washed Delimed Grain (A) | Limed Roan (B) |
0 | 2·88 | 7·03 |
5 | 2·91 | — |
10 | 3·20 | 3·54 |
15 | 4·80 | — |
20 | 4·08 | 4·24 |
25 | 4·29 | — |
30 | 4·85 | 5·19 |
35 | 4·59 | — |
40 | 4·70 | 5·45 |
45 | 4·60 | — |
50 | 4·45 | 5·67 |
55 | 4·42 | — |
60 | — | 5·00 |
The following analyses give the amount of lime in solution before and after the bating of the skins, in grams per litre of filtered puer liquor:—
No. | Bate | Before Goods | After Goods | Increase of CaO |
1 | Fresh puer | 0·108 | 0·62 | 0·512 |
2 | " " | 0·34 | 0·72 | 0·38 |
3 | " " | 0·20 | 0·52 | 0·32 |
4 | " " | 0·19 | 0·49 | 0·30 |
5 | Old puer | 0·54 | 0·84 | 0·30 |
| 6 | Old puer, goods not washed. | 0·98 | 1·38 | 0·40 |
| 7 | French puer shop. | 0·308 | 0·548 | 0·24 |
The limit of the quantity of lime in solution in a normal puer, in the form of calcium salts, appears to be about 1 grm. per litre. If more lime be presented to the liquor, the goods stop coming down. If now fresh dung be added, they will continue to come down, but the quantity of lime in solution does not increase; the excess of lime is precipitated, partly as phosphate and partly as oxalate, in the way stated above.
The type of reaction by which the lime is dissolved is similar to that occurring between ammonium chloride and lime, and is expressed by the formula—
and in the case of the calcium carbonate in the skins—
so that every molecule of lime neutralized, liberates two molecules of ammonia.
Jean states that the production of ammonia progresses regularly during the puering, and that when the free ammonia reaches 0·2 grm. per litre the bate is unfit for further use. But if the excess of ammonia be neutralized by the addition of phosphoric acid, which also precipitates the lime brought in by the skins, the bate may be kept in constant use for some time. Jean found 0·06 grm. NH3 per litre in the puer as made up for use, and, after one lot of skins had been put through, 0·086 grm. per litre; after two lots of skins, 0·135 grm. per litre. In ordinary puer wheels, as used at Trent Bridge, we find—
(a) | (b) | |
| Before goods | 0·0816 | 0·0850 grm. NH3 per litre |
| After " | 0·0833 | 0·0799 |
so that there is little or no difference before and after puering in this case.
Part of the ammonia formed escapes into the air; a further portion unites with the acids which are being formed by the action of bacteria in the bate (see Chapter IV.), and it is also probable that part combines in some way with the calcium salts in the bate.
The actual reactions taking place are of course much more complicated than this simple case, because, instead of ammonia and ammonium chloride, we have a number of complex salts formed by combination of organic acids with substituted ammonia derivatives, such as methylamine, ethylamine, etc. The chief of these are ethylamine and methylamine butyrates, and lactates, and probably propionates, although I have not been able to separate the latter.33
The reaction with the butyrate is represented by the equation—
2C2H5NH2C3H7COOH + CaO
| = 2C2H5NH2 + | C3H7COO | Ca + H2O | |
| C3H7COO |
by which it will be seen that the amine is set free just in the same way as ammonia in the equation on p. 40.
In order to ascertain if the action of the amines was the same as that of salts of ammonia, I prepared the following compounds and tested their action on skins at 37° C., a control piece of the same skin being kept in water at the same temperature. The time in each case was one hour. All the solutions were neutral.
1. Ethylamine lactate: skin swells slightly, not “down” equal to puer.
2. Ethylamine propionate: skin moderately fallen, not equal to puer.
3. Ethylamine butyrate: about the same as Exp. 1.
4. Trimethylamine butyrate: very similar action to Exp. 3 and to ammonium butyrate.
The results correspond very closely with those obtained in previous experiments with various salts of ammonia (see Chapter VI.), and justify us in assuming that in all essential respects the action is similar. The free amines enter into fresh combinations with acids which are produced by bacterial action, and this process goes on until all the nutrient material is exhausted.
The Role of Phosphates in the Bate.—There is no doubt that the phosphates in the bate play an important part, but exactly in what manner they act is not yet known. One of the principal effects is the part they play as “buffers,” in preventing brusque changes of the hydrion concentration during the bating process. This has been pointed out by Soerensen in the case of enzyme reactions. The phosphates in dung are mixtures, which are capable of fixing both acids and bases; and so the small quantities of these bodies, which are produced by the splitting up of the organic matter, are taken up or released as the case may be.
The chemistry of the phosphates is one of the most complicated branches of inorganic chemistry, and, as a consequence, the determination of the constitution of the various phosphates in dung is an extremely difficult matter, and demands a lengthy research. For instance, besides the salts directly derived from the three phosphoric acids, HPO3, H3PO4 and H4P2O7, phosphates exist which are probably derived from hypothetical di-, tri-, or meta-phosphoric acid, nHPO3, and a few salts have been isolated, which are perhaps derived from the hypothetical acids P4O7(OH)6 and P10O19(OH)12 (Watt’s Dict., art. “Phosphates”). Including the double salts, there are more than 16 different calcium salts of phosphoric acids. The normal lime salt Ca3(PO4)2 is very slightly soluble in water, but its solubility is increased by the presence of various organic substances such as exist in dung, and part of the soluble phosphates found in the bate are undoubtedly nothing more than this salt in solution. This fact has been utilized in the manufacture of the artificial bate Erodin (see Chapter VII.).
The phosphoric acid in the puer is partly precipitated by the lime in the skins, and hence diminishes during the bating process. In some cases practically the whole of the phosphoric acid disappears from the solution.
The following experiment will give an idea of the amount of lime precipitated as phosphate. A filtered puer liquor was analysed for lime and phosphoric acid, before and after the skins were passed through. The results were, in grm. per litre—
| Before | After | |
| Soluble lime as CaO | 0·364 | 0·540 |
| P2O5 | 0·424 | 0·328 |
Increase of lime, 0·176 grm.; diminution of P2O5, 0·096 grm. Calculated to calcium phosphate Ca3(PO4)2, this amount of phosphoric acid has combined with 0·114 grm. CaO. Assuming that the ratio of soluble to insoluble lime is the same as given, p. 36, then the lime is distributed as follows:—
| Grm. | ||
| Increase of soluble CaO | 0·176 | |
| " " insol. CaO | 0·210 | |
| Lime as Ca3(PO4)2 | 0·114 | |
| Hence | " " oxalate (?) | 0·096 |
That is, of the lime precipitated, 54 per cent. is phosphate, 46 per cent. oxalate.
In another puer containing before use 0·383 grm. P2O5 per litre, only traces of phosphates were found in solution after goods had been puered in the liquor, and in some analyses by Jean (39) the following figures were obtained:—
Grams of Phosphoric Acid per Litre.
| Fresh dung after four days’ maceration | 0·082 |
| Bate after one pack of skins | 0·036 |
| Bate after two packs of skins | 0·018 |
Although these figures are less than those found at Trent Bridge, they confirm the fact that the soluble phosphates diminish during the bating process.
The phosphates in solution thus diminish during the bating, and are found in the insoluble matter which separates out. A small portion of the lime remaining in the skins is also converted into phosphate by the action of the bate. In an experiment to determine this, a portion of the same skin was taken before and after puering. The pieces were dried, ashed, the ash dissolved in dilute nitric acid, and the phosphates precipitated by ammonium molybdate. In the skin before puering no phosphates were present, but in the skin after puering there was a small amount, though not sufficient to weigh.
The action of ammonium phosphate on the lime in the skin is very small. A skin was treated with a 0·1 per cent. solution of ammonium phosphate at 100° F. for one hour. The CaO in the dry skin was estimated, and found to be—
Per cent. | |
| Before the experiment | 1·93 |
| After " " | 1·45 |
A considerable amount of calcium phosphate was found in the skin after the experiment.
Other chemical compounds existing in the puer, or formed by the action of bacteria (principally B. coli commune, see Chapter IV.), are indol, skatol and a number of aromatic oxyacids, principally para-oxyphenyl-propionic acid, a little para-oxyphenylacetic acid and skatol carbonic acid. In addition, tyrosin, leucin, tryptophan and mercaptans have been separated.34
With these bodies no experiments on skin have been made, so far as I am aware, except with indol and skatol. Kathreiner found that these had a slight reducing action on skin, so that one may say they play some part in the puering.
The action of the bile salts, glycocholate and taurocholate of soda, also needs investigation. These have an indirect effect in the puer, as they favour the development of some species of bacteria (chiefly coli) and hinder the growth of others.
Some action has also been ascribed to sulphuretted hydrogen, but in the puer liquors which I have examined no H2S was found, either before or after the skins were entered.
It will be seen that the ammonia compounds in the bate are not of themselves particularly fitted for the purpose of removing lime35 from the skins, but owing to bacterial action (which we shall treat of in Chapter IV.), acids are produced which combine with the ammonia, and in this way the small quantity of these compounds originally present is continually being regenerated while the bating is in progress. Ammonia is set free by the lime in the skins. It is then neutralized by acids produced by bacteria, and thus acts as a carrier for the acids, and the bate remains in a nearly neutral condition. As the lime increases in the liquid the action of the bacteria diminishes, and finally the alkalinity becomes too great to allow the bacterial or chemical action to proceed further.
It will be noted that the concentration of the active salts in the bate is extremely small. If the amine compounds be assumed to consist of ethylamine butyrate or lactate, the concentration of the solution is approximately 1 grm. per litre; it is important that the concentration of salts should not greatly exceed this amount. I have found by experiment with ammonium chloride solutions, that the best reducing action is provided by a concentration of 0·7 to 1 grm. NH4Cl per litre; if the concentration be raised to 2 or 3 grm. per litre, the skins become “leathery” and do not fall properly. The alkalinity must not be greater than 3–5 c.c. N/10 per 100 c.c. bate, for the bate to work at its best.
Solution of Skin Substance during the Puering.—The determination of the total skin substance dissolved by the puer is best done by Kjeldahl’s method before and after the goods.36
The difference in the total nitrogen found multiplied by 5·6 gives the amount of skin substance dissolved by the bate, assuming the amount of nitrogen in the dry ash-free skin to be 17·8 per cent. If very great accuracy be required, a small correction for nitrogen, brought into solution from the puer itself, is necessary.37 This correction must be ascertained for the particular puer used by actual experiment.
The following figures give the results in grams per litre obtained in the puering of sheep grains. 50 c.c. of the filtered puer liquor are slightly acidified with sulphuric acid, evaporated nearly to dryness and Kjeldahled in the usual way.
Mean | ||||
| Total nitrogen before skins | 0·2632 | 0·2604 | ||
0·2576 | ||||
| " " after " | 0·4928 | 0·4844 | ||
0·4760 | ||||
| Difference | — | 0·2240 |
Equivalent to 1·254 grm. skin substance per litre. This was somewhat over a kilogram of skin substance for the paddle in question, and equal to 1 per cent. of the dry ash-free skin.
As to the differentiation of the dissolved skin substance into albumoses, peptones, monamino acids, diamino acids, ammonia, etc., a modification of Stiasny’s method38 for the examination of soaks and old limes may be used.
The method is based on the fact, discovered by Schiff, that formaldehyde reacts with amino acids, forming methylen-amino acids, which are distinctly acid and allow a sharp titration with phenolphthalein as indicator, while the amino acids themselves react almost neutral. Soerensen has worked out a method on this basis for the determination of different amino acids, and for tracing the course of hydrolysis of albuminous matters.
Instead of using phenolphthalein as an indicator, the electrometric apparatus of Sand (see p. 76) is employed. 50 c.c. of the filtered puer liquor are put into a beaker, the hydrogen electrode is immersed in the liquor, and the potential difference (P.D.) observed; this gives the hydrogen ion concentration of the solution. 10 c.c. of neutral formaldehyde solution (40 per cent.) are added, and the P.D. again observed; it will be found to diminish rapidly, but soon becomes constant, indicating that the reaction is a quick one.
The increase of acidity, as shown by the lowering of the potential difference, is due to the acidity developed by the combination of the formaldehyde with the amino acids forming methylen-amino acids of appreciable hydrion concentration. The amount of such acids is estimated by titrating with N/10 caustic soda solution until the P.D. rises to the same voltage as that originally found. The following figures were found in an experiment:—
Original Puer Liquor | The same After Goods | ||
| π (volts) | 0·61 | 0 | ·69 |
| π after 10 c.c. formalin | 0·53 | 0 | ·54 |
| N (Kjeldahl), grm. per litre | 0·3136 | 0 | ·5936 |
| Increase of N | — | 0 | ·2800 |
| N/10 soda for 50 c.c. to original voltage | 7·0 | 11 | ·6 |
| c.c. increase N/10 soda | — | 4 | ·6 |
| ∴ 1 c.c. N/10 soda = mg N | — | 3 | ·05 |
A preferable method is to add decinormal acid or alkali to the original liquor until the P.D. of 0·69 is reached, at which point the liquor will be neutral to phenolphthalein, and, after adding formaldehyde, to titrate with N/10 soda until the P.D. of 0·69 is again reached.
The factor which connects the amount of decinormal soda required for the titration, after the addition of formaldehyde, with the total nitrogen as determined by Kjeldahl’s method, will afford information as to the extent of the hydrolysis undergone by the proteid matter, in the same manner as Stiasny (loc. cit.), has proposed to differentiate the dissolved proteid matter in lime liquors. As hydrolysis proceeds the percentage of nitrogen in the molecule increases, being at its maximum in the ultimate nitrogenous product ammonia; the factor, therefore, becomes less as hydrolysis becomes more advanced.
| For ammonia 1 c.c. N/10 soda | = | 1 | ·4 mg. N |
| " hydrolized gelatin39 | = | 2 | ·9 " |
| " Witte peptone completely hydrolized39 | = | 3 | ·6 " |
| " lysin | = | 2 | ·8 " |
| " arginin | = | 5 | ·6 " |
| " histidin | = | 4 | ·2 " |
We may conclude that the skin substance dissolved in the puer liquor is hydrolized almost as completely as gelatin is by boiling with sulphuric acid.
I have previously pointed out that dilute acids dissolve a certain amount of skin substance (see p. 157), and in this connexion, Dr. Georges Abt has given me the results of some experiments, on the solubility of skin in various organic acids, which he made in Vienna. Pieces of skin, weighing 40 grm. in the wet state, were allowed to remain for one month in N/10 solutions of the acids. The N was then determined, by Kjeldahl’s method, with the following results, expressed as per cent. of the wet skin dissolved:—
| Per cent. | ||
| Acetic acid dissolved | 0 | ·645 |
| Lactic " " | 2 | ·27 |
| Butyric " " | 0 | ·577 |
| Formic " " | 1 | ·47 |
It will be seen that butyric acid dissolved the least amount of skin, lactic acid dissolving close upon four times as much.
Scud.—A certain amount of skin substance comes away in the “scud.” This is the liquid squeezed out of the skin by the pressure of the scudding knife after puering.
The liquid has the same composition as the puer liquor out of which the goods have been taken, and in addition contains large quantities of pigment granules, wool roots, and some skin substance, which together constitute the so-called “filth” of the skin. Analysis of a scud from English sheep grains showed only 0·164 per cent. N, equivalent to about 1 per cent. skin substance (9·15 grm. per litre). Fat, 7·9 grm. per litre.
Eberle and Krall have recently40 analysed the fatty matter which adheres to the men’s knives in scudding lamb skins for gloving work. They obtained the following results:—
Per cent. | ||
| Water | 29 | ·7 |
| Fat | 42 | ·0 |
| Fatty acids combined with lime | 6 | ·6 |
| Albuminous matter soluble in water | 3 | ·8 |
| Hair and insoluble albuminous bodies | 14 | ·4 |
| Ash (containing 57 per cent. CaO) | 3 | ·5 |
The fat had a—
| Melting-point | 40–44° |
| Saponification number | about 121 |
| Iodine number | 31·6 |
| Hehner value | 91·9 |
| Acid value | 9·3 |
Dr. Fahrion found, in a sample of the fat extracted with ether:—
Per cent. | Iodine number. | |
| Unsaponifiable | 47·6 | 27·3 |
| Fatty acids (sol. in petrol ether) | 39·3 | 30·2 |
| Oxy acids (sol. in ether) | 13·5 | 13·4 |
The figures obtained for the fat therefore agree closely with those for wool fat.
Action of the Bird-Dung Bate.—The depleting action of the pigeon- and hen-dung bate is very similar to that of the puer, or dog-dung bate; but the bating process with these materials, as we have seen (p. 18), is carried out at a lower temperature, and is consequently more prolonged. The principal difference between the two bates appears to be a chemical one, due to the fact that bird dung contains all the urinary products which are present only to a small extent in the dung of mammals. In birds uric acid is the chief stage in nitrogenous katabolism, the mechanism of its formation being a process of synthesis in the liver (Halliburton). Urea is also present in considerable amount, and does not appear to be so easily decomposed as the urea in animal urine.41 As we shall show in the next chapter, urea, and probably also urates, greatly facilitate the permeability of gelatine, and to this fact may be ascribed the more gradual action of bird-dung bates. If we attempt to bate hides with dog-dung, the grain of the hide is found to be attacked and destroyed before the bate has penetrated to the interior of the hide. On the other hand, a bird-dung bate may be used at a temperature of 38° to 40° C. for the puering of skins destined for light leather, but its action is not so favourable as that of the puers.
Composition of Bird Excreta.
— | Pigeon | Hen | Duck | Goose | |||
| Moisture | 58·32 | 56·08 | 60·88 | 46·65 | 77·08 | ||
| Organic matter† | 28·25 | 19·56 | 19·22 | 36·12 | 13·44 | ||
| Phosphates | 2·69 | 2·54 | 4·47 | 3·15 | 0·89 | ||
| Carbonate and sulphate of calcium | 1·75 | 3·08 | 7·85 | 3·01 | 2·94 | ||
| Alkaline salts | 1·99 | 0·82 | 1·09 | 0·32 | |||
| Silica and sand | 7·00 | 17·92 | 6·69 | 10·75 | 5·65 | ||
— | 100·00 | 100·00 | 100·00 | 100·00 | 100·00 | ||
| † Containing nitrogen equal to ammonia | 1·75 | 1·21 | 0·74 | 0·85 | 0·67 | ||
Macadam42 states that pigeon dung is the most concentrated. Hen manure contains the largest proportion of phosphates, and is followed by duck droppings. That of the goose is the least valuable. The preceding table is taken from his paper.
Procter43 quotes the following, as a mean of 40 analyses of pigeon dung by Schulze:—
Per cent. | |
| Water | 21·00 |
| Nitrogen | 2·53 |
| Phosphoric acid | 1·79 |
| Potash | 1·46 |
He remarks that the action of bird dung is more penetrating, but less softening and loosening than that of dog dung, and this effect may be explained by what has been said above.
Unfortunately, far less work has been done on the bird-dung bate than on the puer, and there is a wide field open for research in this direction.
“The most important aspect of any phenomenon, from a mathematical point of view, is that of a measurable quantity.” —Maxwell.
It is impossible in the space of a short chapter to give an adequate explanation of the physical changes taking place during the puering process. An outline only can be given, and perhaps a few signposts to indicate for those interested, in what direction further work can be usefully done. It is obvious that if more be attempted, a treatise on physics and physical chemistry would be required; such a work is beyond the scope of the present volume. It is to be hoped that the whole of the questions dealt with here, including the physical chemistry of skin and of the whole tanning process, will shortly be fully treated by the master hand of our greatest tanning chemist, Professor H. R. Procter.
Since tanning, in the earlier stages, is for the greater part a physical absorption, or colloidal co-precipitating action,44 the physical state of the skin fibre, or condition of the skin before it enters the tanning liquor, is of the greatest importance. In fact, the whole of the operations to which the skin is subjected previous to tanning are directed towards changing its physical state, and the chemical changes undergone are small and principally hydrolytic.
The two most important physical influences over which we have control are pressure and temperature.
The pressure during puering is practically constant, viz. the atmospheric pressure, but, as we shall see later, a diminution of pressure is favourable to falling or depletion. It would, therefore, be interesting to conduct experiments on puering in vacuo, or under reduced pressure.
Before considering the effect of temperature and the changes in the volume of the skin during puering, it will be well to consider the properties of the skin and the puer liquor.
In the skin prepared for tanning, practically all the keratin and epidermis and soluble matters are got rid of, and we have a mass of fibres which are composed of collagen, a proteoïd, which by prolonged boiling with water is converted into gelatin.
Neither the chemical formula nor the molecular weight of collagen is known with certainty, but, from a series of ultimate analyses, the change of collagen into gelatin is represented by Hofmeister45 by the following equations:—
| C102H149N31O38 + H2O | = | C102H151N31O39 |
collagen | gelatin |
from which it will be seen that he assigns a molecular weight of 2416 to collagen, and 2434 to gelatin. Other considerations, however, make it probable that the molecular weight is double the above figures.
Gelatin is a typical colloid, and we may consider that skin is practically a colloid with a structure (a very important point, as we shall see later), but behaving in many ways as gelatin. The puer solution is to a large extent colloidal. Therefore, both the skin and the puer are in the colloidal state. In the typical crystalloid solution of an electrolyte, the dissolved body is separated into its molecules, and to a large extent into individual ions, while, in the colloid state, the units of distribution are either large and often conjugated molecules, or more frequently minute particles composed of many molecules united by cohesive attraction.46 In the case of the skin the molecules are not free to move, but are held in place by the structure of the skin, and the fibres thus act as semi-permeable membranes, with capillary spaces between them, in which water and other fluids are merely held by capillary attraction.
If filtered puer liquor is put into a vessel closed by a membrane of skin, and the whole immersed in clear lime water, the puer solution becomes turbid in a short time, but the outer solution remains clear, showing that the skin is permeable to the lime solution (crystalloid), but not to the puer solution (colloid). It was observed, however, that the acids contained in the puer diffused through the membrane; this was shown by the addition of a few drops of phenolphthalein to the lime solution, when the pink colour disappeared after a short time in the neighbourhood of the skin. These phenomena are due to osmosis. It is the fundamental property of all animal membranes, to allow some substances to pass through them more easily than others. In many cases, such membranes, while freely permeable to water, are practically impermeable to certain substances in solution, and play the part of sieves in directing and controlling the diffusion. In the case of skin the phenomena are complicated by the fact that the skin combines chemically with many substances in solution, and thus we do not always know what part to assign to chemical combination and what to the osmotic phenomena.
Procter has shown (Colloid. chem. Beihefte 1911, ii., pp. 243–284) that, while gelatin is very permeable as such to solutions of acids and salts, there may be formed in the presence of excess of acid a hydrolysable chemical complex of the nature of a salt, in which the gelatin functions as base, and which is probably less permeable to acids and their salts than the neutral gelatin. The conditions would then be similar to those which obtain when solutions of an acid and its salt are separated by a movable membrane, which is permeable for the acid and water but not for the salt solution. From the organized structure of the skin surface, it is unlikely that osmosis takes place between the skin itself and the outer solution, with the two surfaces of the skin as semi-permeable membrane. Osmotic action is most likely to occur in the interior of the skin, between the skin fibres themselves and the interfibrillar spaces. The colloids in the puer solution, which constitute a large proportion of its material, cannot, from their nature, penetrate the skin. This may be shown by the above-mentioned experiment. From this it is reasonable to assume that the lime is not actually dissolved from the interior of the skin by the puer acids, but that solution takes place for the most part after it has diffused out into the puer solution. It is probable, however, that part of the bodies of acidic character present in the puer are capable of penetrating the skin fibre, as has been explained above.
The intensity of the osmotic action of puer upon skin must depend upon the quantity of substances contained in it, to which skin substance acts as an impermeable membrane, and which on that account induces an osmotic pressure between the outer puer solution and the solutions held in the skin fibre. The effect of puering does not necessarily imply the actual expulsion of water from the skin—in fact, well puered skin may quite possibly contain as much water as it did in the swelled condition. The difference consists in the manner in which the water is held in the skin, and its freedom to move from parts which are submitted to pressure. In the swollen skin, the fibres may be conceived as swollen by the water and holding it in the same manner as a gelatin jelly; after puering, the fibres are “fallen,” and the water, hitherto held by them, surrounds them in the liquid form.
The osmotic pressure47 of a solution of concentration c, temperature T, and pressure p, is the difference of pressure exerted on both sides of a semi-permeable membrane in thermodynamic equilibrium, having on the one side the solution under the above condition, and on the other side the pure solvent under the pressure p0 of its own saturated vapour. On this definition the osmotic pressure of a normal solution is over 22 atmospheres, or 330 lb. per sq. in.; and since a saturated lime solution is about 120 normal, its osmotic pressure is about 1·1 atmospheres, or 16 lb. per sq. in.—this represents the force causing the lime to diffuse into water in which the skin is placed. The puer solution being of a colloidal nature, exerts practically no osmotic pressure, and since it contains substances capable of entering into combination with lime, the latter is removed from the surface very quickly. The curve representing the removal of lime by water has been given in Chapter I., p. 6. That for puer is not of such a simple character (see Fig. 7, p. 38), but it will be seen that the greater part of the lime is removed during the first 10 minutes. The curve is plotted for percentage of ash, since the lime is no longer in a caustic condition but in the form of salts. It is remarkable that the percentage of ash, after reaching a minimum, increases considerably. This phenomenon still requires investigation.
Density of Skin.—Coming to the consideration of the volume of skin and its changes during puering, we know that the volume v is the reciprocal of the density, i.e.—
v = 1δ,
and therefore
δ = 1v.
Carini48 has carried out exhaustive experiments on the density of skin during tanning, but, so far as I am aware, little or no work has been done as to the effect of puering on the density of raw skin.
The usual methods of determining density are well known,49 and consist in weighing the body first in air, then in water or other liquid. If m be the weight of the body, and it loses the weight w when weighed in water—
δ = mw.
For experiments on skin, instead of weighing in water it has been found more convenient to use a simple volumenometer, which was devised by Mr. Douglas J. Law (see Coll. 1911, p. 230.)
The apparatus (Fig. 8) consists of the two vessels A and B, connected by means of thick rubber tubing to the burette C, of which the top is enlarged to a bulb. The bottle A, which was specially made for us by Messrs. Townson and Mercer, London, is of about 1 litre capacity, and the wide mouth is closed by a stopper, accurately ground to fit the neck, and extending down to the bottom of the neck. The upper part of the stopper is elongated to a tube, which is closed by the tap G. The vessel A is also fitted with a tap E. The vessel B serves as a reservoir, and is used to adjust the level of the liquid in the burette C by means of the tap at F. To find the volume of a piece of skin, the method of procedure is as follows. The bottle A is filled with water up to the neck, and the stopper D, carefully greased, is inserted. The tap E is then closed, and the burette is filled with water. Then, with the taps G and E open, the bottle A is filled with water up to D by raising the burette. G and E are then closed, and by opening the tap F the level of the water in the burette is adjusted to zero when F is again closed. G and E are then opened again, and, by lowering the burette, the water in A is allowed to fall below the level of the neck of the bottle. E is then shut, the stopper D removed, and the piece of skin is carefully introduced, avoiding air bubbles. The stopper D is then replaced with the tap open, and, by opening E and raising the burette, the water is allowed to come up to the stopper D again. The taps G and E are then closed, and the volume of the piece of skin read off directly from the burette scale. Volumes up to 50 c.c. are measurable as described above, but larger volumes may be measured if a known volume of water is run from the burette into the reservoir before introducing the skin.
By using various solutions in the apparatus instead of pure water, the real swelling or contracting effect of these upon the skin may be observed. The skin is introduced into the bottle A, and the solution adjusted to G, which is then closed; then, by leaving the tap E open, the real swelling or contraction of the skin is measured by the rise or fall of the liquid in the burette.
Petroleum or other liquid may be used instead of water; in some cases, the use of petroleum is more advantageous.
The density of dry gelatin as determined by Lüdeking is 1·412, which is not greatly different from that of skin. Carini gives the following figures for ox-hide:—
| Skin with hair | 1·450 |
| " depilated with lime | 1·425 |
| " depilated with sodium sulphide | 1·441 |
but does not give details how these figures were obtained, and if corrected for ash and fat.
The density of limed and puered sheepskin, determined by drying the skin over sulphuric acid until the weight was constant, then determining the volume in petroleum, gave the following results:—
| Limed skin | 1·2335 | |
| Puered " | 1·2590 | I. |
By determining the volume of the wet skin in the volumenometer, and the per cent. of water on drying the skin, the calculated densities were—
| Limed skin | 1·438 | |
| Puered " | 1·300 | II. |
Correcting for ash and fat, the dry ash-free skins had the densities—
| Limed skin | 1·397 | |
| Puered " | 1·335 | III. |
The density of the wet limed skin was 1·063, but the density calculated from the results (I.) above is 1·0475; from this, it is evident the fibres of the swollen limed skin undergo compression on swelling, or that the water contained in them is in a state of compression, in the same way as gelatin swollen with water occupies a less volume than the sum of the volumes of gelatin and water. Lüdeking50 found for 10 per cent. gelatin jelly δ = 1·069; δ calculated was 1·041. He attributed the whole of the compression to the water, so that 1 c.c. of water in a 10 per cent. gelatin jelly occupied a volume of 0·96069 c.c. See also par. 4 p. 68.
It may be of interest to give the figures used in the preceding lines in tabular form.
| Limed skin | Puered skin | |||
| Density of wet skin | 1 | ·063 | 1 | ·053 |
| Per cent. of water found | 80 | ·63 | 78 | ·2 |
| " " calculated | 79 | ·50 | 79 | ·9 |
| " of ash on wet skin | 1 | ·71 | 1 | ·17 |
| " of fat " " | 0 | ·151 | 2 | ·6 |
| Density of dry ash-free skin substance | 1 | ·3970 | 1 | ·3356 |
In addition to the diminution of the density of the skin by puering, the increase of fat is very marked.
The percentage of water referred to the wet skin may be calculated from the formula
v2 – v11 – v1 × 100
where v1 = specific volume of the dry skin.
v2 = specific volume of the wet skin.
The specific volume of the dry skin (v1) may be calculated from the specific volume of the wet skin (v2), the percentage of water (w) being known by the following formula:—
| v1 = | v2 – w100 | , |
| 1 – w100 |
and by comparing the volume thus obtained with the volume determined by direct experiment, the amount of contraction is ascertained. In the case we have been considering (viz. limed skin with 80 per cent. water)
| v2 calculated | = 0·955 |
| v2 found | = 0·941 |
The specific volume of the dry skin
| v1 calculated | = 0·6955 |
| v1 found | = 0·8112 |
The difference for the dry skin is surprising, and considerably greater than anticipated, but has been verified by a large number of experiments. It may be remarked that the exact determination of the density of skin by drying out pieces is liable to error on account of the non-homogeneity of the pieces. There appears to be some chemical combination between the water and the skin very similar to that between water and alcohol, and although the figures obtained in the above experiments by this method are accurate, it is not certain that they represent the true density. It is possible that the interior of the skin may still contain some moisture, and, in order to arrive at absolute certainty, it would be necessary to powder the skin, and ascertain its density in a finely divided condition.
When this is done, the results obtained are very much more concordant than when pieces of skin are used. The following results were obtained for the density of dry hide powder (as used for analysis by the I.A.L.T.C.):—
| In petroleum ether | 1·2568 |
| In carbon tetrachloride | 1·2570 |
| In alcohol | 1·2580 |
| Mean | 1·2572 |
This is equivalent to a specific volume v1 = 0·7954.
The density of the powdered skin may also be determined in the air volumenometer (Say; Kopp. Kohlrausch Physical Measurements, p. 53).
Swelling and Falling.—The skin, in its living condition on the healthy animal, is the most supple and perfect of coverings, and in producing soft leathers it is the object of the tanner to retain this supple condition. To do this, the swollen fibres must, as we have seen, be got back to their natural state. The phenomena of swelling and depletion of skin have been discussed by Körner,51 and recently Prof. H. R. Procter52 has published a paper which goes into the whole theory of colloidal swelling. In order to understand depletion, which is the opposite of swelling, it is necessary to consider what takes place when a body like skin is swollen. I am indebted to Dr. Th. Körner for the following summary of the phenomena of swelling. There are three types of swelling: 1. Capillary attraction; 2. Endosmose; 3. Molecular imbibition.
The last named is of the greatest importance in tanning. Körner (loc. cit.) enumerates certain principles governing molecular swelling.
1. A body capable of swelling, when put into water, absorbs a definite quantity of the water up to a maximum, which cannot be exceeded. (C. Ludwig.)
2. The maximum of swelling depends upon the chemical composition of the body, on its cohesion and elasticity, and on the temperature and interior pressure of the liquid. (C. Ludwig.)
3. Power of resistance to swelling increases from the exterior to the interior, according to a parabolic law; i.e. the external layers of the body attain the maximum swelling sooner than the internal portions. (L. Mathiessen and A. Schwarz.)
4. The volume of the swollen body is smaller than its original volume, plus that of the liquid absorbed. (Quincke.)
5. Swelling is accompanied by development of heat.53
The production of heat is simply due to the contraction, and not to any chemical phenomenon, such as hydration.54 This explains a fact well known to tanners, viz. that skins swell in cold water and “fall” in warm water. Riecke55 concludes that the degree of swelling, m2/M (where m2 = mass of water absorbed, M = mass of the body swollen), in a space filled with aqueous vapour, unsaturated, is a function of the pressure and temperature.
The velocity of swelling (Pascheles) may be expressed by the formula—
dQdt = (M – Q) K
where M = maximum of swelling, Q = amount of swelling in the time t, and K = a constant.
The differential quotient dQdt gives the velocity for each moment, and it will be seen that the swelling becomes slower and slower as the maximum is approached. Thus, the law of the velocity of swelling is identical with that of the velocity of inversion of cane sugar, itself an application of the law of masses.56
For every process of swelling, the constant K must be determined experimentally from observation of M, and it may be shown that—
K = 1t log MM – Q
whence the value of K may be calculated for each series of determinations.
On the manner in which water is held in swollen colloid bodies, three hypotheses have been put forward.57
1. The hypothesis that colloids have a structure in the form of a honeycomb. (Bütschli.)
2. The water is absorbed at the surface of colloids in a specially condensed form.58
The water forms, with the swollen body, a “solid solution.” (Nägeli.)
Swelling, and its opposite, contraction, are connected with the surface tension between the swelling or contracting bodies and the surrounding solution. With diminished surface tension, the surfaces of contact between the two become greater, i.e. swelling takes place, with simultaneous diminution of the volume of the whole system, and vice versa.
For the absorption phenomena which occur, the following relation holds good: When a substance in solution diminishes the surface tension at the dividing surface, its concentration is increased; it is absorbed. When a substance in solution increases the surface tension at the dividing surface, its concentration is diminished.
In the system water-hide substance the researches of Wiedemann and Lüdkeing have shown that swelling is accompanied by evolution of heat. Since a rise of temperature is favourable to a system formed by absorption of heat, it therefore hinders swelling and vice versa. This is confirmed by practical experience, and most bating operations are conducted at temperatures between 35° and 40° C.
With regard to the influence of pressure on swelling, a similar law holds good as for temperature, which may be expressed as follows. When a chemical system is compressed at constant temperature, its equilibrium is shifted in that direction by which the reaction is accompanied by diminution of volume. Quincke, however, has shown what appears to be a paradox, namely, that swelling is accompanied by diminution of volume, i.e. the swelling substance, plus the water taken up, occupies a smaller space than the sum of the two constituents taken together; therefore, an increase of pressure must be favourable to swelling, and conversely a diminution of pressure is favourable to “falling.”
We must clearly bear in mind that in all cases of swelling, it is the entry of water into the system which is the cause of the swelling, and since puering is a process which acts in a contrary direction, i.e. the skin falls, this means that water is expelled from the system, skin plus water. By direct determination, from 3 to 8 per cent. of water is expelled during puering. Calculations made from density determinations are masked by the fact, above referred to, that the density of the skin changes during puering.
A fallen skin contains less water than when in the swollen condition, but the difference between the percentages of water in the skin in the two states is only relatively small, and obviously insufficient in itself to account for the great difference in the physical properties of the skin in the two conditions. It is evident, therefore, that the state in which the water is held in the two stages must be different. It is reasonable to suppose that, in the case of swollen skin substance, the whole of the capillary cavities of the organized structure of the skin are completely filled with water, which, owing to its incompressible nature, confers upon the skin elastic and unyielding properties which it always possesses in this condition. The water, entering the organized cell structure in the first place by osmotic pressure, is fixed there and confers upon the skin, in part, its own property of incompressibility. It is a well-recognized fact that, during the puering process, large quantities of the skin substance are dissolved by the action of the enzymes in the bate, and it is probable that the finer organized structure is first attacked. The walls of the cells which hold the water are partly destroyed, leaving the skin with, quite possibly, the same amount of water contained in it as a whole, but with the water dispersed at large throughout its interior structure instead of being held in the fibres. The result of this is, that when a small area of the puered skin is subjected to local pressure the water in the portion undergoing compression is free to move into the adjacent portions of the skin, and there is no force acting upon it, except that of capillarity, to cause it to return to that area when the pressure is removed from it. When a portion of a puered skin is compressed, the surrounding part is always swollen to an equivalent amount. That this view is, in the main, correct is supported by an observation of Abt (Paris), that, while cell nuclei may be demonstrated in the layers near the epidermis of an unpuered skin, they are entirely absent from a puered skin, showing that the puer has completely dissolved the nuclear structure of the cells.
The swelling caused by alkalis is of a different nature to the swelling caused by acids. Procter has pointed out59 that alkaline swelling is not repressed by sodium chloride, even when caused by sodium hydrate, but is repressed by sufficient concentration of the hydroxyl ion in the outer solution.
Müller60 points out that jellies containing certain non-electrolytes (glucose, glycerine, alcohol) render the diffusion of soluble matter much slower than pure jellies of the same concentration, but the presence of urea favours the permeability of gelatin and agar jellies. The differences in the velocity of diffusion of various salt solutions through jellies of the same strength, are thus not due altogether to the greater diffusibility of the particular salt solution, but are to be attributed to the influence of the salt on the permeability of the colloidal medium. Limed skin treated with a 1 per cent. solution of urea at 43° C. is depleted or “falls,” but both the solution and the skin remain alkaline.
Přibram61 has recently shown that the swelling and contraction of muscle are to be ascribed to absorption and expulsion of water from the cells of the muscle; the rapid changes taking place, are brought about by changes in the concentration and composition of the liquid components of the muscle. Lactic and phosphoric acids are produced with enormous rapidity, and of relatively high concentration; during a short period these cause instantaneous changes in the swelling of the muscle cells, followed by a return to their original state. For the evidence of this the original paper must be consulted, but it goes to show that the equilibrium of the system—colloids, electrolytes, water—depends on the proportions and quantities of each of these constituents. The phenomena we are considering thus conform to the general law of mass action (Guldberg and Waage).
Influence of Solid Matter in the Bate.—It was first pointed out by Wood (J.S.C.I. 1899, p. 991) that a filtered puer liquor had less action on the skin than an unfiltered liquor containing much finely suspended matter.
On adding an inert solid (kaolin) to the filtered bate, the action was hastened. No doubt much of the colloidal matter, and with it some of the enzymes, are removed by filtration, but nevertheless, the suspended particles in the bate have some effect on the process.
In a most interesting paper, Perrin62 has recently shown, that the granules in suspension in a colloidal liquid function like the invisible molecules of a perfect gas with a molecular weight of 3·3 x 10-9. Such granules are endowed with the molecular (Brownian) movement, and therefore may exert a mechanical effect on the fibres of the skin. Although the mass of the particles is large63 compared with molecular dimensions, they are small enough to penetrate the pores of the skin, and where puering is carried too far, may become deposited beneath the fine hyaline layer, and thus render it cloudy and unsuitable for fine colours.
Hydrogen ion Concentration of Puer Liquors.—It has already been shown, in Chapter II., that fresh puer liquors have a certain acidity (7 c.c. N/10 per 100 c.c.) at the commencement of the operation, but that at the end they are alkaline (3 c.c. N/10 per 100 c.c.). If an artificially acid liquor be made by diluting hydrochloric acid until it shows the same number of c.c. by titration, it will be found far too “strong,” and will swell the skins. This brings us to the consideration of what is meant by the strength of acids.
Procter and Jones64 have drawn attention to the point in their paper on “Acids in Tan Liquors.” As is well known, the ionic theory affirms that degree of acidity depends on the concentration of hydrogen ions, a strongly acid solution being one in which the hydrion concentration is great, an alkaline solution one in which it is extraordinarily minute, and if we adopt pure water as our standard of neutrality, a neutral solution is one in which the hydrion concentration is approximately 10-7 normal. A normal solution of hydrogen ions would contain 1 gram of hydrogen ions per litre; in the case of hydrochloric acid this would equal 1·35 N/1 HCl.
Sand and Law,65 and Wood, Sand and Law,66 have described the mode of estimating the hydrogen ion concentration in tan liquors directly by means of the electrometric method, and this method is especially applicable to the estimation of the hydrogen ion concentration in puer liquors. It can also be used to titrate the liquors, and we have already given some of the results in Chapter II.
The method is based on the theory of Nernst, that the difference of potential between a metal plate and the solution of one of its salts into which the metal is dipping, depends on the osmotic pressure of the free ions of that metal in the solution (in other words, on the concentration of the solution). In the case of hydrogen ions, we use a plate of platinum coated with platinum black and saturated with hydrogen, and the difference of potential depends, therefore, on the concentration of the hydrogen ions in solution.
The accompanying figure (Fig. 9) shows the hydrogen electrode I, and the auxiliary electrode II, drawn to scale,67 on the right, whereas the electrical apparatus is explained diagrammatically on the left.
The principle of the method of measurement consists in connecting the two ends P and Q, of a sliding rheostat to the terminals of a dry cell, D, and balancing the potential-difference to be measured against the potential-difference between one end, P, and the slider, S, by means of a special form of enclosed capillary electrometer, E. The value of this potential-difference is read directly on a delicate voltmeter, V. The connexions, which are found ready-made in the box, have been drawn out, whereas, those to be made by the operator are shown by dotted lines. The steps to be taken by the latter, consist first in taking off the capillary electrometer and manipulating it in such a manner, that on returning it into position the capillary may be partly filled with a thread of mercury and partly with the acid. The terminals, X and Y, marked battery + and −, are connected to a dry cell, and the terminals, Z and U, marked cathode and auxiliary respectively, to the hydrogen and calomel electrode. Very careful insulation of the connexion between the terminal marked auxiliary and the calomel electrode is necessary. The hydrogen is passed through the hydrogen electrode until a constant P.D. between it and the calomel electrode is obtained. This P.D. is measured by moving the slider up and down until no movement of the mercury in the capillary electrometer is observed on depressing the key K marked electrometer.
Fig. 10 is a view of the apparatus as set up for the titration of a puer liquor. H is a cylinder of compressed hydrogen;68 I, the hydrogen electrode, dipping into the beaker C containing the liquor for titration; b, the burette, containing N/10 acid or alkali; II, the auxiliary electrode, the capillary of which is also seen dipping into the beaker C; P, the potentiometer box containing the sliding rheostat S and electrometer E; D, a dry battery. The acid or alkali is run in from the burette until the voltmeter shows 0·69 volts, indicating that the liquid is neutral or has a hydrogen ion concentration of 10-7.
The following table shows some results obtained on puer liquors before and after skins have been put through. p+H has been called by Sorensen the exponent of the hydrogen ion concentration C, and is defined by the equation
p+H = log 1C
i.e. it is equivalent to the logarithm of the reciprocal of the factor of normality of the solution with respect to the hydrogen ions.69 p+H for pure water or a neutral solution is 7, corresponding to 0·69 volts. The measurements were made on filtered puer liquors, using a N/1 KCl calomel electrode as auxiliary, the capillary of the electrode being filled with 3·5N potassium chloride:—
No. | Before goods. | After goods. | ||
Volts. | p+H | Volts. | p+H | |
1 | 0·560 | 4·7 | 0·755 | 8·16 |
2 | 0·585 | 5·16 | — | — |
3 | 0·600 | 5·4 | 0·658 | 6·44 |
4 | 0·595 | 5·35 | 0·710 | 7·35 |
5 | 0·600 | 5·4 | 0·725 | 7·6 |
6 | — | — | 0·770 | 8·35 |
No. 6 was a very old “spent” liquor. The mean hydrogen ion concentration before goods was 0·588 volt, i.e. the concentration was 10-5·32 normal equivalent to 0·00000479 grm. per litre of hydrogen ions. Therefore the value of p+H was 5·32. A solution of hydrochloric acid of the same strength by titration consumed 0·7 c.c. N/1 alkali per 100 c.c. Measured by the electrometric apparatus, it showed 0·410 volt, corresponding to p+H = 2·1, or a hydrion concentration of ·0079N. In other words, the HCl solution has an acidity or strength 1600 times that of the puer liquor.
The mean hydrogen ion concentration of the liquors after goods was 0·000000076 grm. per litre, corresponding to 0·715 volt and p+H = 7·12, i.e. the liquor was alkaline to a slight extent. For comparison saturated lime-water gave a reading of 1·01 volt, corresponding to p+H = 12·5.
The hydrogen ion concentration is of the greatest importance for the proper action of the enzymes in the bate;70 we shall, however, treat of this in Chapter VII.
Conductivity of Puer Liquors.—It was thought of interest to examine the electrical conductivity of puer liquors in actual use, in the hope that the numbers obtained might give some useful indications. It was found that the conductivity increased, as might be expected from the lime going into solution, but the difficulties of the method render it of less use than ordinary chemical analysis. The results of a typical liquor are given here as a record—
Conductivity (K) of liquor before goods
0·00316 1ohm × cm.
Conductivity (K) of liquor after goods
0·00423 1ohm × cm.
The difficulty of expressing the complex reactions of puering numerically is, we have seen, very great, for, as Minot71 says, “with human minds constituted as they actually are, we cannot anticipate that there will ever be a mathematical expression for any organ or even a simple cell, although formulæ will continue to be useful for dealing now and then with isolated details. Nevertheless, the value of graphic methods to every student of science has been immense.”
It has long been my endeavour to express quantitatively the degree to which a skin has fallen. My friend Dr. Sand has suggested that this may be done by subjecting a piece of the skin successively to increasing and then decreasing pressures, and measuring the thickness under each load. Experiments carried out with the apparatus described below show that a limed skin treated in this way is first compressed, and then on releasing the pressure recovers more or less of its former thickness, according to the amount of plumping it has received, i.e. it shows a certain amount of resilience. A well-puered sheep-skin, on the other hand, shows no resilience at all, i.e. on releasing the pressure the whole of the compression persists. In the case of an ox-hide subjected to a bate of hen-dung, a slight recovery takes place on releasing the pressure. This accords with the fact that it will never be possible to puer a thick ox-hide so effectively as a thin sheep-skin. A piece of india-rubber, on the other hand, is completely resilient, i.e. it wholly recovers its thickness on releasing the pressure. The relative thickness of the same skin in the limed and puered conditions under varying loads is also of interest. The process of puering may, as a rule, be taken to reduce a limed skin to between two-thirds and one-half of its thickness in the swollen condition. If both limed and puered skin be then subjected to the same load, the puered skin will at first be compressed very much more than the limed one. This is probably due to the expulsion from it of water, held simply by capillary attraction. On further increasing the load, however, the compression decreases greatly in the case of the puered skin; with both limed and puered skin increase of compression ultimately becomes practically proportional to increase of pressure, and is slightly greater with the former than with the latter.
The table gives representative results obtained on the same sheep-skin (roan) in the limed and in the puered condition. These results are expressed graphically in Fig. 12.
Load per sq. cm. in grm. | Thickness in mm | Δ | |
Limed skin | Puered skin | ||
0 | 3·45 | 1·78 | 1·67 |
20 | 3·43 | 1·58 | 1·85 |
40 | 3·33 | 1·43 | 1·90 |
60 | 3·28 | 1·35 | 1·93 |
80 | 3·22 | 1·23 | 1·99 |
100 | 3·15 | 1·13 | 2·02 |
120 | 3·08 | 1·08 | 2·00 |
140 | 3·03 | 1·05 | 1·98 |
160 | 2·95 | 1·01 | 1·94 |
180 | 2·90 | ·98 | 1·92 |
200 | 2·83 | ·95 | 1·88 |
300 | 2·73 | ·88 | 1·85 |
All off | 2·98 | — | — |
380 | 2·61 | ·82 | 1·79 |
600 | 2·43 | ·76 | 1·67 |
All off | — | ·76 | — |
Δ = difference in thickness of the skin—i.e. compression under the same load.
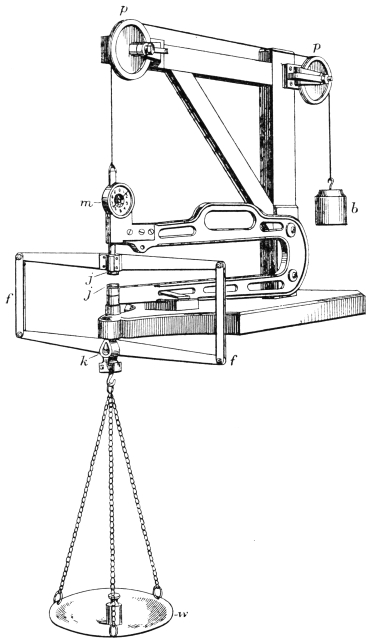
Fig. 11.—Apparatus for Measuring Degree of Falling
m | = micrometer dial. |
JJ | = jaws between which the skin is placed. |
b | = balance-weight attached to cord passing over pulleys p p, to counterbalance weight of frame f f. |
k | = knife edge supporting pan and weights w. |
Fig. 11 shows the apparatus72 that was employed to obtain these results. It consists essentially of a commercial form of micrometer for measuring the thickness of leather. To one of its jaws a pan for weights is attached, by means of the frame f f, in such a manner as to secure a perfectly straight pull. The weights of the frame and pan are counterbalanced in the manner shown by a counterpoise b. The delicacy of measurement may be increased by inserting larger jaws in the form of suitably fashioned disks, but even when this is done the results are to a certain extent vitiated by the rather considerable friction of the micrometer.
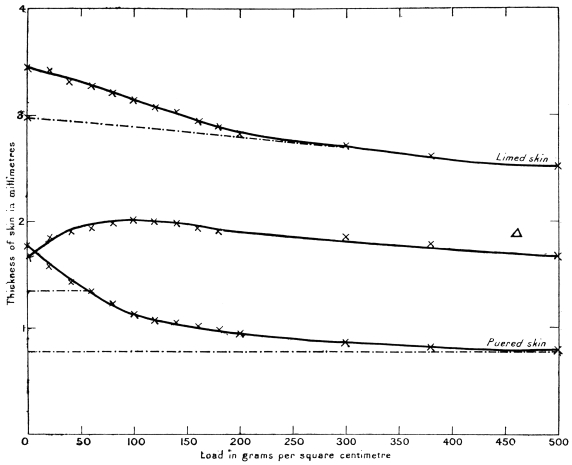
Fig. 12.—Curve obtained by Measuring Apparatus.
(The dotted lines show the pressure on taking off the load.)
An apparatus free from this fault is shown in Fig. 13. It consists essentially of a counterbalanced lever A, to which the upper jaw J is rigidly attached. By means of a sliding weight W, any desired load, from zero upwards, may be put on this jaw. The lever carries a very delicate spirit level, which allows it to be set accurately horizontal in every experiment. The lower jaw is movable vertically between parallel guides, and its position is controlled by the screw-wheel S which bears a divided circle on its circumference. The position of this wheel, and therefore of the lower jaw, may be accurately read on the vernier v. In every experiment it is adjusted so as to make the upper lever accurately horizontal.
“Omne vivum ex ovo.”—Harvey (1578–1675).
When a drop of liquid from a puer wheel in use is examined under the microscope73 with 112 o.i. objective, it is seen to be swarming with bacteria.
The majority are short rods (bacilli), but other forms, cocci and spirilli, are seen in lesser numbers. Most of these bacteria move briskly in the liquid; as the temperature of the slide sinks, their movements become slower, and finally cease. The illustration, Fig. 14, shows the various forms of bacteria observed by the author in puer liquors × 1000 diam.
The living bacteria are best examined in a drop culture in the following manner. A clean cover-glass, of the proper thickness for the objective to be used, is laid upon a black glass plate. With a platinum loop, previously heated to redness in the flame, a drop of sterile physiological salt solution (0·6 to 0·75 per cent.) or sterile broth is placed in the centre of the cover-glass. With a platinum needle a minute quantity of the puer liquor is stirred into the drop. A slide with a depression in the centre is taken, the edge of the depression painted round with vaseline, and pressed over the cover-glass, so that the drop is exactly central. If the whole be now turned smartly over, the drop will hang central in the hollow space.
If the ring of vaseline is continuous, and the cover well pressed down, the drop is preserved from evaporation, and the bacteria may be examined in their natural condition—best on the edge of the drop.
For illuminating the drop culture, the concave mirror is used, and a small diaphragm without condenser; whereas, for stained preparations, the flat mirror is used in conjunction with the Abbe condenser.
If the cover-glass be carefully removed, and dried under a bell glass, the culture may be preserved in a dry condition, or may be stained and mounted.74
If the dried preparation is on a cover-glass, it should be held in the fingers (prepared side upwards), and passed slowly three times through the flame of a Bunsen burner. By holding the preparation in this way, the exact temperature for proper fixation is obtained.
A drop of fuchsin stain, or gentian violet, is allowed to remain on the preparation for five minutes; wash off the superfluous dye with water, and examine, either in the wet state or after drying and mounting in balsam.
For a detailed account of the technique of staining and mounting, the following works may be consulted:—
Methods and Formulæ. P. W. Squire. (Churchill.)
Taschenbuch für den bakteriologischen Praktikanten. Dr. Rudolf Abel. (Stubers-Verlag, Würzburg.)
Technique Microbiologique. Nicolle and Remlinger. (Octave Doin, Paris.)
Practical Bacteriology. Kanthack and Drysdale. (Macmillan, London.)
The recent researches of Tissier and Metchnikoff have shown that the flora of the intestines, both of men and animals, consist very largely of anaerobic bacteria. These have been overlooked in previous researches, owing to imperfect means of studying this class of organisms. Indeed, in one work on the microbes of the alimentary canal of the dog, no mention was made of them, whereas they are all very active.
Most of these organisms, and the new methods by which they have been isolated, are fully described in a new work entitled “Les Anærobies,” by M. Jungano and A. Distaso, of the Pasteur Institute, Paris.75
The following bacteria have up to the present been isolated from dung (mostly dog dung), and studied in pure cultures:—
| 1. | Micrococcus ureae (Cohn). (Pasteur.) | ||
| 2. | " fulvus (Cohn). | ||
| 3. | " prodigiosus. | ||
| 4. | " ureae liquefaciens. | ||
| 5. | Bacterium sulphureum. | ||
| 6. | " coli commune. (Fig. 15.) | ||
| 7. | " coli anindolicum. | ||
| 8. | Bacterium coli anaerogenes. | ||
| 9. | " furfuris α (Wood). (Fig. 30.) | ||
| 10. | " furfuris β (Wood). (Fig. 31.) | ||
| 11. | Bacillus fluorescens putridus. | ||
| 12. | " " " liquefaciens. | ||
| 13. | " subtilis. | ||
| 14. | " saprogenes (Herfeld), three varieties. | ||
| 15. | " butyricus (Hueppe). (Fig. 23.) | ||
| 16. | " putrificus. (Fig. 19.) | ||
| 17. | " pyocyaneus. | ||
| 18. | " janthinus. | ||
| 19. | " coprogenes fœtidus. | ||
| 20. | " pyogenes fœtidus (a variety of B. coli). | ||
| 21. | " zenkeri. | ||
| 22. | " magnus. | ||
| 23. | " spinosus. | ||
| 24. | " liquefaciens (Eisenberg, Frankland). | ||
| 25. | " amylobacter (Van Tieghem). | ||
| 26. | " acidi paralactici. | ||
| 27. | " I. | Isolated from horse manure by Severin, Centr. Bl. f. Bakt. (2), i., 97. | |
| 28. | " II. | ||
| 29. | " III. | ||
| 30. | " from horse dung (anaerobic) Severin, Centr. Bl. f. Bakt. (2) iii., 708. | ||
| 31. | " from horse dung (anaerobic), No. 3, ditto. | ||
| 32. | " oedematis maligni (Vibrion Septique, Pasteur). | ||
| 33. | " mesentericus vulgatus. | ||
| 34. | " lactis aerogenes. | ||
| 35. | " cavicida (Brieger). | ||
| 36. | " albuminis (Bienstock). | ||
| 37. | " Bienstockii. | ||
| 38. | " tenuis. | ||
| 39. | " enteritidis sporogenes (Klein). | ||
| 40. | " lactis acidi (Ankerschmid, 1905). | ||
| 41. | " megatherium. | ||
| 42. | " cadaveris sporogenes (Klein) said to be identical with No. 16. | ||
| 43. | " thermophilus. (Houston). | ||
| 44. | " a. from puer. See p. 162. | ||
| 45. | " b. " " " | ||
| 47. | Bacillus mycoides. | ||
| 48–61. | 14 species isolated from dog and pigeon dung by Prof. H. Becker. Zeit. f. Offentlich. Chemie. Heft xxiii. Jahrgang X. p. 447, includes B. erodiens (Fig. 16). | ||
| 62. | Sarcina fimentaria (Lehmann and Neumann). | ||
| 63. | Streptococcus from sewage. (Houston.) | ||
| 64. | " brevis. | ||
| 65. | " longus. | ||
| 66. | " pyogenes. | ||
| 67. | " liquefaciens coli. (Gamgee Phys. Chem. 2.) | ||
| 68. | Streptothrix from stable manure. (Severin, 6.) | ||
| 69. | Spirillum serpens (Kutscher). | ||
| 70. | " tenue " | ||
| 71. | " undula " | ||
| 72. | " volutans " (Figs. 24 and 25). | ||
| 73. | " from pig dung. Smith, Centr. Bl. f. Bakt. 16, (1), 124 | ||
| 74–76. | Vibrio, three species isolated by Kutscher. | ||
| 77. | Clostridium butyricum (Prazmowski), said to be identical with No. 25. | ||
| 78. | Streptococcus faecalis. Sidney Martin, 37 and 38; Ann. Rep. Loc. Gov. Board, 1907–9; Nature, March 3, 1910, p. 22. | ||
| 79. | Bacillus bifidus. | ||
| 80. | " perfringens. | ||
| 81. | " bifermentans. | ||
| 82. | " funduliformis (Veillon). | ||
| 83. | " capillosus. | ||
| 84. | " sporogenes. | ||
| 85. | " ventriosus. | ||
| 86. | " rodella III. | ||
| 87. | Staphylococcus parvulus. | ||
| 88. | Diplococcus orbiculus. | ||
| 89. | Coccobacillus preacutus. | ||
| 90. | Coccobacillus oviformis. | ||
| 91. | Bacillus faecalis alkaligenes (Petruschky). | ||
| (79–90 are anaerobic bacteria, described and figured by Jungano and Distaso.) | |||
It will be surmised from the above list, to which additions are still being made, that the flora of the intestines is pretty extensive, and, consequently, the study of the part played by the various species of bacteria is a long and difficult one.
The methods of isolating these bacteria, and the compositions of the media employed, would demand a treatise on bacteriology; but, for general purposes, a good liquid medium for the cultivation of puer bacteria is a gelatin peptone broth, made by digesting 10 grm. gelatin with 6 12 grm. 80 per cent. lactic acid in 100 c.c. water under pressure for three hours, neutralizing with ammonia, adding 1 grm. potassium phosphate, making up to 1000 c.c., and filtering. A sterile infusion of fresh dung may be used, but it is troublesome to prepare and not easy to get uniform in strength or composition. The culture liquids are left slightly alkaline, an alkalinity equal to 0·0636 per cent. Na2CO3 or 12 c.c. N/1 soda per litre. The amount of alkali may be increased to 0·15 per cent. Na2CO3 without affecting the growth of the bacteria. Of solid media, 10 per cent. nutrient gelatin, or in summer 15 per cent., is good if used at temperatures below 25° C. For higher temperatures, up to 39° and 40°, nutrient agar is required. The best nutrient gelatin for general work is made according to Klein’s formula.76 For media in general, a most useful compendium is Abel’s Taschenbuch.
The number of bacteria in fresh fæces varies greatly, but is of the order of 10,000,000 per grm. of dry matter, capable of developing in nutrient gelatin. Of this number, about 100,000 are spore-bearing organisms. This estimate applies to healthy animals; in a diseased condition, the numbers vary enormously.
Dr. A. C. Houston found in raw London sewage from 3,000,000 to 9,000,000 microbes per c.c., of which more than one-tenth were gelatin-liquefying organisms. There were only about 300 spores of aerobic bacteria, about 100,000 B. coli, 100 B. enteritidis sporogenes, and streptococci, in one gram of fæces.
With the object of ascertaining the effect of the various species of bacteria contained in the dung upon skins, a large number have been isolated, and the effect of pure cultures in different media has been tried upon skin.77 A number of the results have been published in the Journal of the Society of Chemical Industry. Professor H. Becker, who has done a great deal of this part of the work, is of opinion that the principal organisms concerned in the bating exist in the dog’s intestines, and belong to the group of coli bacteria. These are very widely distributed bacteria, and are found in the large intestines of mammals, and, as a consequence, in almost all soils, and in the mud of rivers and lakes. The principal variety is B. coli commune.
Lortet found it, along with other organisms, in the mud of the Lake of Geneva, at a spot where the water was chemically very pure. Dr. A. C. Houston, the bacteriologist of the Metropolitan Water Board, enumerates sixteen varieties of this organism, 80 per cent. of which produced acid and gas in lactose-peptone cultures, indol in peptone-water cultures, and when grown in milk produced acid and clot. The bacterium (Fig. 15) resembles that of typhoid fever, and has frequently been mistaken for it. It is, however, much more resistent to destructive influences. It is a short bacillus, possessing flagellae, by which it moves more or less rapidly.
B. coli forms short rods 0·8 µ wide, 1 to 3 µ long. It moves somewhat slowly by means of flagellæ, which may be demonstrated by staining with Loffler’s method.78 It grows equally well in absence or presence of air, that is, it is a facultative anaerobe. Although it will grow at room temperature, the optimum growth is at 37° C. In plate cultures the appearance of the colonies below the surface of the gelatin is quite different from that of the surface colonies. The former are small round colonies, about the size of a pin head; the latter spread into a whitish iridescent film, with irregular edges.
B. coli does not liquefy the gelatin. When grown in nutrient solutions containing sugars, it produces much acid, and at the same time gases are given off, consisting of CO2 and hydrogen. If the growth in this solution be allowed to continue a secondary fermentation ensues, and the culture eventually becomes alkaline.
Indol is produced by B. coli, and may be demonstrated by adding to 10 c.c. of the culture, 1 c.c. of a 150 per cent. solution of pure potassium nitrite; then adding a few drops of concentrated sulphuric acid, when, if indol be present, a red coloration (nitroso-indol) is produced. This bacterium reduces nitrates to nitrites. Cultivated in a 1 per cent. solution of peptone, to which 110000 per cent. of potassium nitrate has been added, after four hours at 37° C., the presence of nitrite may be shown; after the growth has continued for seventeen hours, the nitrite is further reduced to ammonia. Among other products of B. coli, Harden found lactic, formic, acetic, and succinic acids, ethyl-alcohol, CO2 and hydrogen.
In Germany, W. Lembke and H. Becker have specially investigated the bacterial flora of the dog’s intestines. Lembke, in 189679 cultivated the bacteria from the fæces of the dog, fed in various ways—bread, meat, and fat diet—and found B. coli constantly present, although the form of the individuals, as well as the colonies, and the intensity of the indol and gas formation, showed great variations.
The other species of bacteria present varied with the kind of food; this has a great influence on the flora of the intestines, which was found to be very different when the dogs were fed on bread to what it was when they were fed on meat.
Lembke describes two other species of bacteria closely resembling B. coli, one of which he calls B. coli anindolicum, which, as the name implies, gives no indol reaction; the other, B. coli anaerogenes, is non-motile, possesses no flagellæ, and differs from B. coli by the absence of gas production in the fermentation of sugars.
Besides B. coli, there are several species of bacteria which liquefy gelatin, and a number of facultative organisms, whose presence is more or less accidental. By changing the food, and introducing with it quantities of foreign organisms, the composition of the intestinal flora may be changed. By introducing for a considerable period B. coli anindolicum, Lembke succeeded in entirely suppressing B. coli commune. On returning to normal feeding, the foreign organisms in some cases entirely disappeared.
The researches of Dr. H. Becker80 were applied more directly to the use of bacterial cultures for the bating of skins, and to the elucidation of the bacterial action of dog-dung infusions. He isolated 54 varieties of bacteria from dog-dung, and tried the action of pure cultures of many of them on a skin.
A list of the various bacteria isolated by Becker is given in tabular form on pp. 98–101.
Professor Becker’s Bacterium No. 12, which he has named Bacillus erodiens (Fig. 16), is undoubtedly a variety of B. coli, but has a more rapid motion, and does not coagulate milk, although it renders it somewhat thick. Cultivated in broth it gives off much gas, consisting of 12 per cent. carbon dioxide, 85 per cent. hydrogen, 3 per cent. oxygen. If glucose be added, the quantity of carbon dioxide rises to 40 per cent., and acid is produced. The most rapid growth is at 37° C., and at this temperature a broth culture has a distinct reducing action on skin. According to the medium in which it is grown, it produces acid or alkali, and thus comes under the heading of mixed bacteria. In sugar81 solutions acid is produced, and in proteid solutions ammonia compounds, indol, and evil-smelling gases are given off. Thus, by varying the medium, the effect produced may also be varied.
B. erodiens does not secrete any tryptic enzymes, hence its action on the skin is to be attributed either to an intracellular enzyme, or to its chemical products, which, being secreted in situ, have a more favourable and powerful action than if merely added to the bating liquid. It was for this reason that I proposed to use a mixed culture of bacteria, especially bacteria from the sweating process (see p. 105), which secrete a mild form of proteolytic ferment, capable of dissolving the more easily soluble portion of the skin fibres (or certain constituents), but not capable of attacking the hyaline layer.
(continuation of table to right)
The practical difficulty is to keep such cultures uniform during propagation, and so far this has prevented their introduction in practice. Similar difficulties have influenced the use of pure cultures of yeast in the brewing of English beers, although the use of a single species of yeast is common in the low fermentation breweries on the Continent.
I found in studying the bacteria of dog dung, that the species existing in the fresh dung, which developed in ordinary plate cultures, appeared to belong to four or five species only, mostly bacilli. At the end of two or three weeks, the original species had given place to others, mostly cocci, in a very similar way to the change which takes place in putrefaction. In fact, many of the organisms are identical with those which cause putrefaction. It will be seen, therefore, that no single species produces the complex chemical and physiological changes which take place, or the bodies necessary for the bating of skin, as some observers have supposed; but the various species succeed one another as the medium changes its reaction and composition, until finally the organic portion is resolved into the simplest bodies such as carbon dioxide, ammonia, and hydrogen. There is thus a moment when the dung is at its best so far as the bating action is concerned, and this moment is due to the vital activity of bacteria, and consequently varies according to the temperature and some other influences (electrical condition of the atmosphere, etc.). One may say it is at its best at about fifteen days in summer, and one month or more in winter. Puer which has been dried, is not so powerful in its action as that which is immediately made into a paste with water. It appears to lose its “nature,” partly owing to irreversible dehydration processes, and partly because some of the bacteria are killed. Plate cultures on agar from fresh puer (Fig. 17), and from a puer wheel in use (Fig. 18), show the number of bacteria in the puer wheel to be much greater than in the fresh puer. Whence it is evident, that the bacteria continue to develop in the puer and to produce their various products, enzymes, etc. We have already considered the action of the chemical products, and in Chapter V. we propose to discuss the action of enzymes.
Pigeon-Dung Bate as used for Hides.—The bacteria contained in the intestines of birds and in bird dung have not been studied to the same extent as those of mammals, so that it is not possible to give anything but a meagre account of them. A microscopical examination of fresh pigeon dung, collected on a sterile Petri dish, showed debris of food, cellulose, etc., among the debris, a large number of dumb-bell bacteria (b) (Fig. 20), and a few motile pairs (c); no bacilli were seen. Cells of a saccharomyces (a) were also observed. From this pigeon dung attenuations were made by a modification of Soyka’s method,82 and from the fourth attenuation a plate culture was made in ordinary nutrient gelatin. The colonies from this plate were principally of two varieties (both non-liquefying organisms), corresponding to the bacteria observed in the original dung. Large cultures were made in a Carlsberg vessel, as described in Chapter VI., and the effect of these cultures tried upon skin. No particular reducing effect was obtained.
A microscopical examination of a bating pit used for kips, showed an extraordinary mixture of bacteria, bacilli, vibrios, and monads; some comparatively large dumb-bell shaped bacteria, very motile, were present. The difference between the bating liquor and the fresh dung was very marked, especially in the variety of species present. Cultures made from several colonies isolated from the above bate, in a nutrient liquid, consisting of 10 litres water, and 20 grm. gelatin, peptonized by heating under pressure with 10 c.c. sulphuric acid, afterwards neutralizing with ammonia, and adding the soluble matter from 200 grm. bone-meal, had no action on skin.
It would be unsafe to say from these two experiments that the bacterial effect of the pigeon-dung bate is negligible, but we may assume that it is different and not so great as with the dog-dung bate or puer.
A complete research as to the various species of bacteria developing in the bird-dung bate is necessary before this question can be answered.
General Considerations on the Growth of Bacteria in Various Media.—Since the publication of Further notes on the action of the dung bate (Chapter VI.), I have found that the bating organisms grow better in the special medium, when it is neutralized with ammonia, than when it is neutralized with sodium carbonate, i.e. the presence of organic ammonium salts is more favourable to the growth of the bacteria than the corresponding sodium salts.
I also found that bacteria obtained from other sources than dung, viz. from the roots of wool just beginning to “slip” in a sweating stove, were equally effective in causing the skin to fall. Now these bacteria produce ammonia, and it seems clear that they are essential to the chemical part of the process. They also produce proteoclastic enzymes, which act upon the skin fibre (see chapter on Enzyme Action). The products of the bacteria depend very much upon the composition of the nutrient medium. Many organisms grown in media containing sugar or other carbohydrates produce acids, but, grown in proteids free from sugars, they produce alkaline compounds. Villon (“The Leather Industry,” 1901, p. 408) describes a bacterium which he considers to be the special micro-organism concerned in the depilation of skins, which resembles Bacillus d (Wood) (Fig. 21), but he does not describe the appearance of the cultures; he states, however (p. 410), that this is the only bacterium which can develop in the limes,83 and that it is the cause of the unhairing in this case also. Since the production of ammonia in limes is known to be due to bacterial action, it is very probable that this bacterium, which is ubiquitous, is also of use in the bate, and a research in this direction would be interesting.84
Some of the fermentations taking place in the dung come under the heading of putrefactive processes (see p. 116). Tyrosin is formed in considerable quantities during putrefactive fermentation, but is soon further decomposed, according to Nencki, with formation of indol, CO2 and hydrogen. Leucin gives valerianic acid, ammonia, CO2 and hydrogen; nitrogenous bodies of the aromatic series are also produced.
Bacillus ureaæ, B. prodigiosus, and B. fluorescens putridus, evolve trimethylamine (Herfeldt), and, as the writer has shown, this amine has an important action in the puering process. In combination with organic acids, it removes lime from the skin, and in addition it favours the growth of bacteria, such as bacillus d and e (Figs. 21 and 22) and B. coli.
The albumens and peptones of the dung are pretty well decomposed and absorbed before evacuation; the bacteria subsequently split up the amido acids into fatty acids and ammonia. The fatty acids are then decomposed generally in the form of the calcium salts, in the manner shown in the table (p. 108), for which I am indebted to Dr. E. Herfeldt, of Bonn.
We have already treated of the action of these various products in Chapter II., but it will be seen from what has been said in the present chapter that the chemistry and bacteriology of the puer overlap, and that it is difficult, if not impossible, to separate them entirely. The bacteria are continually manufacturing chemical compounds, and decomposing others.
In this respect it is interesting and instructive to note that Nencki, in his classical work “The Chemical Mechanism of Putrefaction,”85 considers the processes by which the putrefaction of proteids is brought about by bacteria, to be analogous to those taking place by melting the bodies with potash, and he holds the view that in the hydration processes brought about by bacteria, the water plays the same part as the potash.
| No. | Fermenting Substance. | Cause of Fermentation. | Fermentation Product. | Authors. |
|---|---|---|---|---|
| 1 | Calcium formate | Bacteria from sewer slime. | Calcium carbonate, CO2 and H | Hoppe-Seyler, Archiv f. d. g. Physiol. xii. |
| 2 | Calcium acetate | " " | Calcium carbonate, CO2 and CH4 | " " |
| 3 | Calcium lactate Undergoes four different fermentations | Thin bacillus | 1. Propionic acid, and, as by-products, acetic acid, succinic acid and alcohol. | Fitz, nine papers in the “Berichte der Deutsch. Chem. Gesellschaft,” 1876–1884. |
| Other species of bacteria: short aerobic, butyric bacteria (Fitz). | 2. Propionic acid and valerianic acid. | |||
| 3. Butyric acid and propionic acid. | ||||
| 4. Butyric acid, according to Pasteur (Comptes rend. 1861) | ||||
| 4 | Calcium malate | Bacteria (not described). Thin bacilli. | 1. Chief product, propionic acid; and, as by-product, acetic acid. | Schützenberger, “Fermentation,” 1876. |
| 2. Chief product, succinic acid; and, as by-product, some acetic acid. | ||||
| 3. Butyric acid and H. | ||||
| Bacteria | 4. Lactic acid and CO2. | |||
| 5 | Calcium tartrate | Different species of bacteria. | 1. Chief product, propionic acid; by-product, acetic acid. | " " |
| 2. Butyric acid. | ||||
| 3. Chief product, calcium acetate; by-products, ethyl alcohol, butyric and succinic acids. | ||||
| 6 | Calcium citrate | Small, thin bacilli | Acetic acid in large quantities, along with small quantities of ethyl alcohol and succinic acid. | Fitz. |
| 7 | Calcium glycerate | Micrococci | 1. Calcium acetate, along with small quantities of succinic acid and ethyl alcohol. | " " |
| Medium-sized bacilli | 2. Formic acid, with some methyl alcohol and acetic acid. |
Nencki explains, for example, the metamorphosis of leucin by putrefaction in this way: The bacteria decompose the water into hydrogen and hydroxyl, which act upon the leucin as follows:—
| CH3 | CH — CH2 — | CH | | — COOH + | OH H | = NH3 | |
| CH3 | ||||||
| Leucin | NH2 | |||||
| + OH — CH2 (CH2)4 COOH (oxycaproic acid) | ||||||
The resulting oxycaproic acid is then split up by the second water molecule into methylenglycol and valerianic acid:—
| OH | + | OH H | = CH2 (OH)2 + CH3 (CH2)3 COOH |
| | | |||
| CH2 | |||
| | | |||
| (CH2)4 | |||
| | | |||
| COOH |
The methylenglycol, which changes into formaldehyde and water, is now split up into CO2 and hydrogen, as it would be by melting with caustic alkali.
| H | + | H.OH | = CO | OH | + 2H2 | |||
| C | O | |||||||
| H | H.OH | OH |
As we shall see in the chapter on the action of enzymes, the phenomena are of a catalytic nature. Any urea present is decomposed, by the direct action of micrococcus ureæ, into ammonium carbonate and ammonium carbamate, so that it does not play any part in the bating process as usually carried out with dung which has been kept for some time, but the ammonia produced plays an important part in the chemical action of the bate, as we have already seen.
If, however, dung containing the urinary products be used in a fresh condition, the urea has indirectly a very important influence on the bating, as it favours the permeability of the skin fibre. (See p. 72.)
The fermentation of the cellulose in the dung has not been studied from the bating standpoint, but it is well known that it is fermented by various species of bacteria, which have been grouped together under the generic name of Amylobacter.
Deherain and Gayon first showed that the solution and fermentation of cellulose in the form of dead vegetable matter, which had previously been observed, also took place in dung. Van Tieghem, in 1879, showed that the solution of cellulose is caused by bacteria, whose properties correspond with those described by him as Amylobacter. Tappeiner was able to ferment cellulose by mixed cultures of bacteria from the intestines of oxen—in neutral solution, CO2, methane, H2S, aldehyde, butyric acid, and acetic acid, were all recognized. In alkaline solutions, the principal products were CO2 and hydrogen, together with the same by-products as before.
From the researches of Van Sennis, in 1890, it seems pretty certain that the fermentation of cellulose is due to the symbiotic action of at least two different organisms The decomposition of the cellulose may be explained by considering that first a sugar-like carbohydrate is formed by hydrolysis, and that this is then split up into equal volumes of CO2 and CH4. It may be noted that the fermentation is anaerobic, and no doubt, so far as bating is concerned, the chief products are the organic acids produced, principally butyric and acetic acids. Van Sennis nearly always found Clostridium butyricum associated with this fermentation.86
Another group of organisms which have some influence in the bating process, are the class called by Beijerinck, Granulobacter. They produce butyric acid, and this acid, combining with the ammonia compounds of the dung, forms salts which undoubtedly exert an effect on the lime in the skins, though its action on the fibre is, perhaps, not so great as the compounds of lactic and propionic acids.
The most common butyric ferment is the old Clostridium butyricum, now known as B. butyricus, (Prazmowsky), which is anaerobic. It forms spindle-shaped spores, hence the name Clostridium (from κλωστηρ, a spindle). Another species (Fig. 23), found in milk by Hueppe (1884), is aerobic, and ferments lactic acid and its salts to butyric acid, CO2, and hydrogen; it appears to correspond with Granulobacter polymyxa of Beijerinck.
Oxalic acid is known to be produced by some bacteria and the moulds Penicillium and Sclerotinia, and in the white rot of the turnip it is produced by Pseudomonas; it is also produced by some saccharomycetes, such as B. Hansenii.87 There is reason to believe that its production plays a part in the bate, as we have already mentioned in Chapter II., but the organisms producing it and their mode of action still remain to be investigated.
There are, of course, a large number of putrefactive bacteria in the puer, among these B. putrificus (Fig. 19), isolated by Bienstock; it is a spore-bearing anaerobic bacillus, and is interesting as specially attacking fibrin. Now fibrin is extremely resistant to the action of most putrefactive bacteria, and it is very probable that specific organisms ferment the different albuminous compounds, in the same way that the different carbohydrates are each decomposed by specific ferments.
Very interesting are the various forms of spirilla met with in dung; Figs. 24 and 25 show Spirillum volutans in the unstained condition, and also stained to show the flagellæ. It will be noted that the appearance is so different that, to an inexperienced observer, they might be taken for different species. The rôle played by these organisms still requires investigation.
I have pointed out previously the importance of the nutrient medium, or substratum, in which the bacteria grow, on the species surviving. In it one can see on a small scale the Darwinian process of natural selection. There is a great struggle for existence between the various species, and the circumstances determining the survival of this or that organism are extremely complicated, and we are yet very much in the dark as to the action of the various chemical compounds contained in the puer, so that it is unsafe to neglect even those which are present in only small amounts. Very minute quantities of certain bodies, almost too small for detection by chemical means, are sufficient to cause large differences in the growth of certain organisms. For instance, Raulin found that the addition of a trace of zinc to his nutrient liquids increased the crop of the mould Aspergillus niger more than four times the weight of a crop grown in the same liquid free from zinc.
If we inoculate a nutrient material with a pure culture of bacteria, and the medium is not exactly adjusted to the needs of the particular organism, it will not thrive, and will speedily be overgrown by some other species obtaining access from the air. This fact very much discounts the use of pure cultures of bacteria which have been proposed for bating, although in the case of erodin, where the medium has been adjusted to suit the organism, considerable success has been attained. The whole of the enzymes and chemical compounds essential for a perfect bate, are not present in the dung when it leaves the animal’s body, but these compounds are produced by the continued action of the intestinal bacteria and other organisms which obtain access from the air. The production of the enzymes depends, too, upon the composition of the nutrient medium, since this exerts a selective influence on the species of bacteria obtaining access to it. Just as in the spontaneous souring of milk numerous bacteria have free access to it, yet the lactic ferment is generally so pure that it may be, and is, used as a pure culture on a large scale in the manufacture of lactic acid.
Coming to the action of the bacteria on the skin fibres, from the work of Abt and Stiasny,88 we may conclude that the substance of the conjunctive fibres is less profoundly decomposed by bacterial fermentation than by the action of lime. The latter dissolves about 2 per cent. of skin substance from a fresh skin, whereas a puer acting normally dissolves about 1 per cent.
The nuclein of the skin fibres appears to be all removed by the puer, since Abt confirms the fact that no nuclei can be seen under the microscope in a puered skin. The actual solution of the skin substance is brought about by enzymes of a tryptic character. (See Chapter V.)
While the main lines of the bacteriology of the dung bate are now pretty well known and understood, it will be seen that much work still remains to be done as to details, and this principally with the anaerobic bacteria of the dung, which have been studied by few investigators.89 I have suggested90 that such a research might well be undertaken by the bacteriological laboratories of our Leather Industries Schools in Leeds and London.
Moulds and Putrefaction.—In view of the fact that moulds are of frequent occurrence on dog dung, a brief mention of them is necessary. So far as our present knowledge goes the researches of Van Tieghem, De Bary, Rankin, Marshall Ward, V. H. Blackman and others indicate that their action on the essential bating constituents of the dung is a destructive one. They grow usually on acid media, and in so doing break down the acids present into simple inorganic bodies, such as CO2 and water, utilizing the carbon and nitrogen for their own growth. Although these fungi secrete almost all varieties of enzymes (Bourquelot), yet we have no evidence that any of the enzymes contained in dog dung are from this source. In the usual case of dung preserved in pits or casks, the upper surface only becomes mouldy, since moulds require a free supply of oxygen. The mycelium penetrates but a very little way into the body of the dung, and cannot therefore effect any decomposition, except of the surface layer. The dung exposed to the action of the mould is generally a bad colour, and is rejected as unsuitable for puering.
The following species have been noted and classified as growing on dog dung, though probably not all of them are specific.
1. Pilaira dimidiata (Grove).
2. Mucor caninus (a variety of Mucor mucedo).
3. Circinella simplex (Van Tieghem).
4. Pilobolus crystallinus (also on cow-dung).
Certain myxobacteria are found on dung, among these Chondromyces, described as long ago as 1857 by Berkeley, and at that time included among the Hyphomycetes. It was rediscovered in 1892 by Thaxton, and owing to his researches the whole class of myxomycetes is now generally considered as a division of Bacteria. Another myxomycete, Polyangium primigenum (Quehl), forming a red fructification on dog dung, is figured in the Encyclopædia Britannica, XI. edition, vol. 3, p. 163.
The following abstract gives some account of putrefaction, and may be of use in conjunction with the account of the bacteriology of the bate which has been given. Since it was written Dr. G. Abt (see Bibliography 51) has also given a very full description of putrefactive processes as affecting leather manufacture. The subject is still occupying the attention of a large number of bacteriologists, and we may expect more light to be thrown on the whole question during the next few years.
Abstract of Paper on Recent Advances in the Bacteriology of Putrefaction. Read before the Nottingham section of the Society of Chemical Industry, January 24, 1906.91
To those who have to do with the manufacture of leather, the changes which take place in the skin from the time it leaves the animal are of the utmost interest. The most important of these changes is the natural process of decomposition known as putrefaction.
Putrefaction may be defined as the decomposition of nitrogenous organic matter by living organisms, accompanied by the evolution of malodorous gases. The study of it may be divided into two parts—(1) the biological, (2) the chemical. The first concerns the organisms which break down the proteid molecule either directly or by means of enzymes; the second that of the different products of the action of these organisms. It is extremely difficult to separate these two studies.
Dr. Sims Woodhead (59) gives a concise account of the earliest researches on the organisms causing putrefaction by Leeuwenhoek (1692), Plenciz of Vienna, Müller of Copenhagen (1786), Needham (1749), Spallanzani (1769), Schwann (1837), Schroeder and Van Dusch (1854), Tyndall (1870), Lister (1878). These names show that the history of putrefaction proceeds parallel with the evolution of the microscope and the development of the comparatively recent science of bacteriology. I propose to-night briefly to carry it up to the present day.
I need scarcely say that putrefaction is not a specific fermentation like alcoholic or acetic fermentation, but that it is extremely complex. In any putrefying matter, such as gelatin or albumin, a large number of different species of bacteria may be observed as well as monads and infusoria, and in some cases moulds, all of which take part in the process. The first stage is a process of oxidation in the presence of air, in which ærobic bacteria use up the oxygen present and only simple inorganic compounds are formed, carbon dioxide, nitrates and sulphates; this part of the process is generally without odour. The second stage, or true putrefaction, takes place in the absence of oxygen by anærobic bacteria, and is a process of reduction. It has been shown that there are no bacteria in healthy tissues, and if a muscle or any organ is taken from an animal under antiseptic conditions it may be preserved indefinitely in a sterile vessel to which filtered air has free access. Solid matter is usually liquefied by organisms like B. liquefaciens magnus, which are invariably present in the air, and which prepare the way for more specifically putrefactive bacteria, such as Proteus vulgaris and B. putrificus, but if one observes a number of putrefactions of the same kind of matter under natural conditions, scarcely any two follow the same course. The modern study of putrefaction dates from Hauser (58), who, in 1885, isolated from putrefying flesh the three organisms—Proteus vulgaris, P. mirabilis, and P. zenkeri. He studied the action of these in pure cultures, and came to the following conclusions:—
That Bacterium termo (Ehr.) is not a single definite species; various forms and stages of other organisms have been described under this name. The various species of Proteus go through a wide range of forms during their development in which cocci, short and long rods, thread forms, vibrios, spirilli, and spirochætæ occur. Under special nutritive conditions Proteus goes through a swarm stage, in which condition it is capable of moving over the surface and in the solid gelatin. The Proteus bacteria are facultatively anærobic, they all cause putrefaction; P. vulgaris and P. mirabilis are the commonest and most active of all putrefactive bacteria. They do not secrete an unorganised ferment, but decompose albuminous bodies by direct action. They also produce a powerful poison, of which small quantities injected into animals produce septicæmia.
Tito Carbone (60) found amongst the products of P. vulgaris, choline, ethylenediamine, gadinine, and trimethylamine. Macé (61), criticising Hauser’s work, considers the cocci form of Proteus to be spores. Bienstock (62) believes the rôle of the Proteus group somewhat doubtful. He discovered (1884) another widely distributed putrefactive organism, which he called Bacillus putrificus; it is a spore bearing, drumstick shaped bacillus found in fæces; it is anærobic and specially attacks fibrin. Now fibrin is extremely resistent to the action of most putrefactive bacteria, and it is very probable that specific organisms ferment the different albuminous compounds in the same way that the different carbohydrates are each decomposed by specific ferments. A certain number of species of bacteria are able to decompose both carbohydrates and proteids. Tissier and Martelly (70) call these mixed ferments, and divide them further into two groups (1), mixed proteolytic ferments, including B. perfringens, B. bifermentans sporogenes, Staphylococcus albus, Micrococcus flavus liquefaciens, Proteus vulgaris, this group decompose albumin by means of tryptic enzymes. (2) Mixed peptolytic ferments are only able to attack the albumin when it has undergone a preliminary decomposition. This group comprises B. coli, B. filiformis, Streptococcus pyogenes, Diplococcus griseus non liquefaciens.
The second class of bacteria are those which are without action on carbohydrates, and only attack proteids; these consist of the true proteolytic bacteria B. putrificus, and B. putidus gracilis, and the peptolytic bacteria, Diplococcus magnus anaerobius and Proteus zenkeri, which can only decompose peptones.
These authors state that B. putrificus is always present in putrefying albumin, but always accompanied by facultative ærobes which favour the growth and development of the special putrefactive bacteria.
In the putrefaction of meat the reaction is first acid owing to the action of the mixed ferments on the sugars present. In the next stage ammonia is formed by the tryptic enzymes secreted by the ærobic bacteria, and so the anærobic organisms are enabled to develop. We can thus understand how it is that putrefaction proceeds more rapidly the more mixed ferments there are present, although these were formerly supposed to hinder putrefaction from taking place.
When meat is exposed to air it is first attacked by the mixed ferments, Micrococcus flavus liquefaciens, Staphylococcus, Bacillus coli, Bacillus filiformis, Streptococcus and Diplococcus, and becomes acid; at the same time, the presence of decomposition products of albumin may be detected, proteoses, amidoacids, amines and ammonia; the latter quickly neutralise the acids, and in three to four days the meat is alkaline, and has a faint putrid smell. Bacillus perfringens and Bacillus bifermentans sporogenes now make their appearance; the latter of these organisms produces amines, amido-acids and ammonia. In this stage the simple anærobic ferments are able to begin their work, and real putrefaction sets in; as this proceeds, the mixed ferments gradually disappear, and finally the only organisms remaining are Bacillus putrificus, Bacillus putidus gracilis, and Diplococcus griseus non liquefaciens.
Another organism, which appears to play an important part in the decomposition of animal bodies, is described by Klein (63); he found that in bodies, which had been buried from three to six weeks, bacteria such as B. coli and B. proteus had almost disappeared, and an anærobic bacterium, which he calls B. cadaveris sporogenes, was very active. It is a motile bacillus 2–4 µ long, with flagellæ all over its surface. Spores are formed at the rounded ends, giving it a drumstick form. It coagulates milk, the clot gradually dissolving. It grows on all the usual nutritive media, but only under strictly anærobic conditions.
In a paper, entitled “Fermentation in the Leather Industry,”92 I gave a short account of the progress of putrefaction as it takes place in the animal skin, and also described some of the organisms I had observed in putrefying skin. A small piece of skin was placed in water and allowed to stand at room temperature. During the first two days there was little change, but on the third day a number of swiftly moving darting monads made their appearance. Some of these were propelled by flagellæ, but a few had assumed amœboid forms. A slowly moving bacillus consisting of a long straight rod, apparently broken up into cells exactly like the Vibrio subtilis, illustrated in the “Micrographic Dictionary,” was observed, accompanied by some species of spirillum. Higher organisms present were a Paramœcium and a colourless transparent piece of protoplasm, shaped like a dumb-bell, with a slow rotating motion. On the fifth day the number of vibrios and spirilli had greatly increased, some with a swifter motion than others. There were also many large infusoria present; one of a peculiar double form, which appeared to be a development of the dumb-bell shaped piece of protoplasm seen on the third day. On the seventh day the most striking feature was the great increase in the number of vibrios; the field of the microscope was crowded; masses of the bacilli could be seen clustered round small particles of the disintegrating skin as if feeding upon it; there were more infusoria, many of them short, boat-shaped monads, with a trembling motion, refracting light strongly; these evidently accompany the putrefaction bacteria, and assist in the final disintegration. On the ninth day the piece of skin was entirely dissolved.
Procter calls attention to the relative putrescibility of the different constituents of skin, and especially to the rapid putrescence of the lymph and serum. So far as I know, this part of the subject has not been studied at all thoroughly, and there is a considerable field open to workers in our research laboratories.
Pure fat is not decomposed by bacteria, but if albuminous matter is present, the fat is split up by several species of bacteria and moulds. Schreiber (73) has shown that the presence of oxygen is necessary. As this subject scarcely comes within the category of putrefaction, I refer you to Schreiber’s paper, and also to an important paper by Otto Rahn (74) recently published.
In the putrefaction of vegetable matter the cellulose is attacked by specific organisms, which have been thoroughly investigated by Omeliansky (75). He has shown that the fermentation of cellulose is an anærobic process, caused by two species of bacteria belonging to the class of butyric ferments. Morphologically the organisms closely resemble one another, but one of them decomposes the cellulose with evolution of hydrogen, the other with evolution of methane; in both cases considerable amounts of acetic acid and normal butyric acid are produced.
I have previously stated that monads and infusoria take part in the process of putrefaction, but I do not know that their action has been studied in the same way as that of bacteria. The life history and morphology of some of these monads was studied in 1871 to 1875 by Dallinger and Drysdale (76). These authors, in their researches into the life history of the monads found in a putrefying infusion of cod’s head, came to the conclusion that “bacteria are not the only or even (in the end) the chief organic agents of putrefaction, for most certainly in the later stages of a disintegration of dead organic matter the most active agents are a large variety of flagellate monads.”
Dallinger cultivated some of the monads in Cohn’s fluid, and found that they lived and multiplied in it. Their spores were killed at a temperature of 250° F. There is a big field of research open in this direction.
The consideration of the chemical aspect of putrefaction is a vast subject, and would demand a special treatise. I shall only call your attention to one or two points of interest.
Taking the simpler bodies first, sulphuretted hydrogen is formed in putrefying liquids in two ways: (1) by reduction of the sulphates in the liquid by an anærobic organism Spirillum desulfuricans; (2) by bacteria capable of growing in the presence of oxygen such as B. coli commune and B. lactis ærogenes, which ferment glucoses with formation of lævorotatory lactic acid and evolution of CO2 and hydrogen, and if at the same time the material contains albumin or sulphur, H2S is given off; these organisms are incapable of reducing sulphates. Beijerinck (64) has investigated this process, and found a variety of different forms intermediate between the two above-mentioned, but all possessing the same characteristics so far as their chemical action is concerned, so that they may be classed as one order, which he calls Aerobacter.
Stich (65) found phosphorus pentoxide in the residue from the putrefaction of casein, nuclein, lecithin, and protagon; and in the putrefaction of certain organs of animals and plants, gases containing phosphorus are evolved. The nucleic acid of yeast yielded phosphoric acid along with hypoxanthin and xanthin.
Vitali (66) found in the putrefaction of muscle, which had been freed from sugars and fat, that some alcohol was produced. He considers that a hexose is split off from the albumin in a similar manner to the splitting off of a fermentable sugar from the glucoproteids (compounds of simple proteins with carbohydrates). The formation of alcohol in the putrefaction of muscle occurs in the alkaline stage. Thus alcoholic fermentation is caused not alone by saccharomyces, but also by certain putrefactive bacteria.
Lermer (77) finds that the putrefaction of barley resembles butyric fermentation. An analysis of the gases given off during the later stages of the process gave the following result: nitrogen, 58·88; hydrogen, 37·43; methane, 3·15. In the residue from the putrefaction he found acetic, butyric and valerianic acids, but not caproic or caprilic acid. In the normal steeping process employed for barley the gases given off consisted almost entirely of carbon dioxide and nitrogen. This observation is interesting to compare with the evolution of nitrogen in the fermentation of bran shown by Wood and Willcox.93
The action of putrefactive bacteria has been found capable of transforming hexoses into pentoses. Salkowski and Neuberg (78) inoculated a solution of d-glukuronic acid with putrefying meat, and showed that it was changed into l-xylose with evolution of CO2 according to the following formula:—
COH(CHOH)4COOH = CO2 + COH[CHOH]3CH2OH.
This is an interesting fact, especially as, according to Neuberg, the pentose contained in animal nucleo-proteids is l-xylose.
I wish to express my indebtedness to Dr. Alfred Koch’s “Jahresbericht über Gärungs-organismen” for some of the abstracts.
The following is a list of putrefactive bacteria which have been studied in pure cultures:—
1. Proteus Vulgaris (Hauser). 2. Proteus mirabilis. 3. Proteus Zenckeri. 4. Bacillus Oedematis maligni (Kerry, Nencki, Bovet). 5. Bacillus Chauvæi = B. sarcophyematos bovis. 6. B. Liquefaciens magnus. 7. B. spinosus. 8. B. putrificus (Bienstock). 9. B. pseudo œdematicus (Liborius). 10. B. enteritidis sporogenes (Klein). 11. B. tetani. 12. Clostridium fœtidum. 13. B. cadaveris sporogenes (Klein). 14. Spirillum desulfuricans (Beijerinck). 15. B. coli commune. 16. B. lactis ærogenes. 17. B. fermentationis cellulosæ. 18. Micrococcus flavus liquefaciens (Flügge). 19. Diplococcus griseus non liquefaciens (n. sp.). 20. Streptococcus pyogenes. 21. Staphylococcus pyogenes albus. 22. Bacillus filiformis ærobius (n. sp.). 23. Diplococcus magnus anærobius (n. sp.). 24. Bacillus putidus gracilis (n. sp.). 25. B. perfringens (Frankel). 26. B. bifermentans sporogenes (n. sp.).
Moulds taking part in putrefaction, principally of fruit and vegetable matter:—
1. Penicillium glaucum. 2. Mucor mucedo. 3. Mucor piriformis (Fischer, possibly identical with 2). 4. Mucor stolonifer (Ehrenberg). 5. Botrytis cinerea (Pers). 6. Mucor racemosus (Fres). 7. Monilia fructigena (Pers). 8. Fusarium putrefaciens (Osterwalder). 9. Cephalothecium roseum.
“The living organism is enabled by the use of enzymes to bring about, under ordinary conditions of temperature and moderate concentrations of acid or alkali, many chemical reactions which would otherwise require a high temperature or powerful reagents.”—W. M. Bayliss.
In a recent work,94 Dr. Bayliss defines enzymes as the “catalysts produced by living organisms.” A catalyst is a body which greatly accelerates the rate of reaction in chemical processes, without apparently taking part in the process. For instance, peroxide of hydrogen is decomposed into oxygen and water by mere contact with finely divided platinum, while the latter remains unaltered in the process. In this case the platinum black is the catalyst.95
In natural processes the best known type of an enzyme is diastase (amylase), the enzyme contained in malt, and which enables the malt to convert starch into dextrin and sugar (maltose). It is capable of transforming more than 2000 times its own weight into sugar, which fact is quite sufficient to show that its action differs from that of an ordinary chemical reaction. Another enzyme, sucrase, according to O’Sullivan and Thompson, will hydrolyze 100,000 times its weight of cane sugar to invert sugar. Rennet will coagulate 250,000 times its own weight of casein in milk. The list of enzymes grows longer almost daily, as some new one is separated having a specific action, until one is almost led to believe that the mechanism of life itself, as manifested in the cell, is due to enzymes.
It has been found that enzymes act very much in the same way as inorganic catalysers. As an example, the velocity of the reaction of invertase (the enzyme of yeast which hydrolyses cane sugar to grape sugar) has been compared with the same hydrolysis brought about by heating a solution of cane sugar with a mineral acid. In both cases the reaction is in accordance with the law of mass action (Guldberg and Waage) that the amount of sugar transformed will decrease as less remains to be transformed. In the diagram (Fig. 26) the curve A is for invertase (Jas. O’Sullivan, “Journ. Inst. of Brewing,” vol. v. p. 168); curve B is for the hydrolysis by acid (Wilhelmy), from which it will be seen that the manner in which the hydrolysis proceeds is practically the same in both cases.96
In the case of fermentation by the living organism, the fermentation rises rapidly, and then gradually slows down and comes to an end before the whole of the fermentable matter is used up; the curve, therefore, is of a hyperbolic character, C in the diagram, which represents, in a general way, the fermentation of glucose by B. furfuris. The ordinates represent the amount of acid produced by the bacteria. The time in this case would be 176 more nearly represented by hours instead of minutes on the abscissa.
The mathematical expression for the velocity of the reaction is
dxdt = k(a − x)
where
a = original concentration of solution
x = the quantity transformed in time t
k = coefficient of velocity of the reaction
By integrating the above equation, it may be shown that
1t log aa – x = k
For the experiment of J. O’Sullivan with invertase, k has a mean value of 0·0013; for Wilhelmy’s experiment with acid, k has a mean value of 0·001377.
Enzymes are produced by the living cell, with other secretions, and as a consequence are found in all plants and animals. Certain organs, however, produce enzymes in large quantities, or appear to be specially set apart for their production. In plants, the seeds are the chief seat of enzyme activity; in animals, certain glands, such as the salivary glands and the pancreas. The mucous membrane of the stomach and intestines also secrete enormous quantities.
The production of enzymes by bacteria was observed by Wortmann in 1882. It has been found that the secretion of the enzymes depends upon the composition of the nutrient medium in which the bacteria are grown. For instance, Pfeffer found that the secretion of diastase by Bacillus megatherium depended upon the amount of cane sugar in the nutrient medium. The cane sugar checked the secretion of the diastase, and the same effect was observed in the case of the common mould Penicillium glaucum. In Bacillus mesentericus vulgatus, diastase has been found to exist side by side with four other enzymes. Passini,97 in studying the putrefactive anaerobic bacteria of the normal human intestines, succeeded in separating from B. putrificus a proteolytic enzyme filtered free from bacteria, and which caused proteolysis in media which were too acid to permit of the bacteria growing. The enzyme easily dissolved, without previous neutralization, the coagulated casein caused in milk by old coli cultures. Acid gelatin media were also liquefied by the enzyme.
From the evidence we have at present it seems probable that every variety of enzyme, hydrolysing, oxidizing, ammoniacal, etc., can be produced by bacteria.
The following useful classification of enzymes is due to Effront.98
A more recent classification based on chemical properties is that of Kossel and Dakin.99 They divide ferments into two classes:—
(1) Oxylytic ferments capable of breaking the O-link by which the radicals are held together in fats and carbohydrates.
(2) Imino-lytic ferments, including the amino-lytic ferments which act on the amino groups of urea.
Group 2 is sub-divided into—
(a) Trypsin and erepsin, which separate the imide NH from the neighbouring carbonyl CO.
(b) Arginase, which separates off urea from arginin.
Although it is somewhat doubtful whether the enzymes contained in dog dung are of glandular origin,100 it is quite certain that other enzymes are secreted by bacteria developing in the dung while it is kept prior to being used for puering. These enzymes may be separated by the following method. About 150 c.c. of puer is well mixed with an equal quantity of glycerin, and allowed to stand for seven days. It is then filtered through paper by means of a pump, and yields a clear filtrate of a deep golden-brown colour; the filtrate is poured in a thin stream into a tall vessel containing about 1500 c.c. of 98 per cent. alcohol. A copious flocculent precipitate of the albuminous matter and enzymes is thrown down, the solution is filtered and the precipitate washed on the filter with absolute alcohol, and then dried over sulphuric acid in vacuo. The resulting powder is, of course, a mixture of all the albumins in solution, and probably only a small portion of it consists of the pure enzymes. We have merely succeeded by this method in concentrating them. The property of albuminous bodies in the act of coagulation to carry down soluble matter is well known, and this also renders the preparation of any pure proteid extremely difficult. It may be mentioned here that recent evidence goes to show that enzymes are not of a proteid nature, since, by repeated purification, the proteid matter may be almost entirely got rid of, while the activity of the residue containing the enzyme becomes considerably greater.
Krawkov’s method of preparing diastase from saliva (Green, p. 46) may also be mentioned, as it is of general application. It consists in salting out the enzymes, by saturating the clear solution (in this case saliva diluted with an equal volume of water) with neutral ammonium sulphate. The precipitate which is caused by the saturation is collected on a filter, and washed for a short time with strong alcohol. It is then allowed to stand under absolute alcohol for one or two days, and finally dried at 30° C. On extraction with water it yields a solution which is strongly diastatic, and which gives no proteid reactions.
There are several other methods for the preparation and purification of enzymes, but up to the present it may safely be stated that no one has succeeded in preparing an enzyme in a state of purity.
In considering the mechanism by which enzymes act, it must be remembered in the first place that they are colloids, and, as such, will form absorption compounds with the substrate, or body, upon which they are acting. It is difficult to understand how an enzyme can exert an action on the substrate, unless it enters into some kind of combination with it, although this may be only a temporary one. The action of some enzymes has been found to be due to extremely small amounts of certain metals, e.g. in the case of the oxidizing enzyme laccase, the metal is manganese. In the purest samples of this enzyme prepared, 0·16 per cent. Mn was found, and it has been supposed by some observers that the whole of the action of the enzyme may be attributed to the physical state of the manganese which it contains. According to this hypothesis,101 the active part of the enzyme is the ion Mn. This ion may exist in the solution in two conditions, differing by the electric charge which they carry. One of them Mn++ carries two positive charges, the other Mn+++ carries three. In the first phase, Mn++ is transformed into Mn+++ by absorbing the charge of one ion of hydrogen (H+), and two hydrogen ions thus discharged, in the nascent state, unite with the oxygen dissolved in the liquid to form water.
2Mn++ + 2H+ + 0 = 2Mn+++ + H2O
In the second phase the ion Mn+++ with three charges will be transformed into the ion with two charges, by decomposing a molecule of water, of which the nascent oxygen will attach itself to the oxidizable body R yielding the oxide RO.
2Mn+++ + H2O + R = 2Mn++ + 2H+ + RO
and the same cycle of operations will begin again, and continue indefinitely.
In the above illustration the enzyme action was an oxidizing one. In the case of puering it is a hydrolytic action, in other words a molecule of water is added to the skin substance. The fibre or some portion of it is converted first into proteoses, and finally into peptones, and simpler bodies. The active metal in these cases appears to be calcium, but by what mechanism it brings about the hydrolysis is at present unknown.
Pozerski102 found that the pancreatic juice, which is secreted after injections of certain sera (anti-pancreatic action), and as a consequence has no pancreatic action, contains no calcium, but pancreatic juice secreted under the influence of pilo-carpin is more or less rich in calcium, and its proteolytic action increases about equally with the amount of calcium contained in it. The same probably holds good for the intestinal juice.
Victor Henri has shown that the power of metals in the colloidal state to bring about these catalytic actions varies with the metal employed, and is in inverse ratio to the size of the particle. There is a very interesting and wide field of research open here in order to determine the conditions under which the various metals act. To this end the ashes from the purest enzyme preparations might be studied, and methods devised for producing these metals in the colloidal state, for it seems evident that it is the state of the body acting which gives it the properties observed, and not its chemical properties in the usual sense.
The enzymes contained in dog dung which are effective in the puering process belong to several groups, principally the proteolytic and lipolytic groups, but indirectly enzymes of the first group (see p. 132), (fermenting carbohydrates) and of the fifth group (fermenting urea) also play some part by decomposing various compounds (e.g. cellulose and urea) contained in the puer.
The action of certain enzymes from the animal body upon skin has been tried by the author.103 Those selected were pepsin and pancreatin,104 as being most likely to be present in dung. Pepsin only acts in presence of HCl. Two portions of the same skin were taken, one of them was treated with a 1 per cent. solution of pepsin, acidified with 0·2 per cent. of hydrochloric acid; the other in a bate liquor of dogs’ dung (puer), both at a temperature of 40°C. At the end of one hour the skin in the pepsin solution was considerably “fallen,” but that in the manure solution was bated nearly away, i.e. the greater part of it was dissolved. A 1 per cent. solution of pancreatin (Mercks) was found to act far more rapidly than pepsin; 1·5 per cent. of chloroform was added to the solution, to prevent the development of bacteria. The skin was reduced, but had not the peculiar touch of a puered skin. As will be shown later, this was found to be due to the absence of any chemical action upon the lime salts in the skin, and consequently it felt “limey.” This action took place in the puered skin, but not in the skin treated with pancreatin alone.
W. J. Salomon (19) has also attributed the activity of the bate to pepsin and pancreatin, but he does not give any proof of the presence of these ferments in the bate.
Since it is practically impossible up to the present, to separate these enzymes from the dung in a state of purity, the method described on p. 134 was employed.
The enzymes prepared in this way consist of a mixture of all the enzymes present in the dung, the amount obtained from 1000 grm. of dung being about 4 grm. The product had a slight diastatic action upon starch; 0·5 grm. in 100 c.c. of water at 35° C. was found to have a very considerable reducing action upon skin, and when combined with the amine compounds prepared from the dung, the action was more powerful, and more rapid than with puer. Limed skin was puered in thirty minutes in this solution to a perfect condition, in the absence of bacteria, and with no evil smelling compounds. The reaction of the solution at the beginning of the experiment was faintly alkaline; at the end of the experiment it was considerably alkaline.
This experiment proves that the action of the dung is a complex one, due to the combined action of enzymes and chemical compounds upon the skin. These compounds, which are principally amines, and salts of amino-acids, probably assist the enzymes, and at the same time act upon the lime remaining in the skin from the previous liming process. Whether a skin, which has never been submitted to the liming operation, could be bated by enzymes alone, without the addition of amines, has not, so far as I know, been tried, but it is highly probable that this would be the case.
In order to compare the action of the enzymes prepared from dung with that of the enzymes produced by bacteria, a mixed culture of bacteria from puer in dextrose gelatin, after seven days’ growth, was taken. 200 c.c. of this was mixed with 200 c.c. of dilute alcohol (65 per cent.), and well shaken: gelatin and albuminoid bodies are by this means precipitated. The liquid was filtered and poured into eight times its volume of 98 per cent. alcohol. The precipitate which came down was washed with absolute alcohol, and dried in vacuo. The enzymes thus obtained were re-dissolved in water, and the former experiment with the skin repeated with this solution, with the addition of the amines. The skin was brought down in exactly the same way as before, showing conclusively that it is the enzymes produced by bacteria, acting in conjunction with the amines, which bate the skin. It would seem that the action of the enzymes is aided by the presence of amine compounds, in addition to the chemical action which these latter have upon the skin. The action is interdependent, i.e. bacterial action alone is insufficient, and chemical action alone is insufficient, the true bating action being a combination of the two.
There seems little doubt that it is the enzymes which dissolve the skin substance, or rather certain parts of the intercellular substance of the fibres, and the compound of this substance with lime. The action is a digestive one, and may be compared, as we have shown, to that of the digestive ferment of the pancreas.
This fact has been made use of in the artificial bate “Oropon” (see Chapter VII.), in which extract of pancreas is combined with ammonium chloride, and some inert material.
The effect of puering on the fatty matter in the skin is well known. The fat and grease are partially emulsified, and set free, so that they can be removed by scudding. This action is a most important one, and one in which artificial bates have hitherto been wanting. The emulsification of the fats is brought about by means of an enzyme either identical with, or closely resembling, lipase.105 This enzyme is found in the pancreatic juice, and in the seeds of many plants. It brings about the emulsification of the fat by saponifying a portion of it, i.e. the fat is split up into glycerin106 and a fatty acid, according to the equation—
C3H5(C18H35O2)3 + 3H2O
Stearin.
= C3H5(OH)3 + 3(C18H35OHO)
Glycerin. Stearic Acid.
Lipase was one of the first enzymes in which the reversibility of the reaction was shown, i.e. it is not only capable of hydrolysing a fat, but also of causing the formation of one by the combination of the fatty acid and glycerin.107 This explains why the reaction of such an enzyme is never complete. An equilibrium is reached just as with ordinary reversible chemical reactions like the precipitation of magnesium hydroxide by ammonia.
Lecithin, and possibly other fatty compounds, are known108 to be important auxiliaries in the ferment-like actions produced by toxins; cholesterol has a similar effect, and as this body is a constituent of dung, it may play some part in puering. Here, again, is a further problem awaiting investigation.
Loevenhart has shown that the bile salts, sodium cholate, and sodium glycocholate, greatly increase the activity of lipase, and Magnus found that synthetic bile salts have the same effect; such bodies are known as co-enzymes. I have shown that bile itself is not favourable to the bating action, but the bile salts, by stimulating the action of the fat splitting enzymes, are probably essential to the full action of the dung bate.
Another enzyme which may be of importance in puering is erepsin, the enzyme of the intestinal juice, which is responsible for the completion of the digestive process. The pancreatic enzymes act upon the peptones produced by the pepsin of the stomach, splitting them up into simpler compounds, while the erepsin acts further upon these products. It dissociates albumoses and peptones into amino acids, taking as it were the last traces of nutriment from the food passing through the intestine. It acts best in alkaline solutions.
This ferment is very widely distributed in the animal kingdom, and occurs in other organs and tissues besides the intestines. The quantity of erepsin in the fresh fæces must be considerable, since a dog secretes from 400 to 500 c.c. of intestinal juice per day. It remains to be shown whether this retains its properties after excretion, and for how long.
A most important point in connexion with the activity of enzymes is the reaction of the medium, i.e. its acidity or alkalinity, or, more strictly speaking, its hydrogen ion concentration. A very slight increase or decrease of the acidity or alkalinity of the liquor will diminish the rate of action of the enzyme by a large amount, and in some cases cause the action to cease. In all cases enzymes have an optimum acidity or alkalinity; in other words, for every enzyme there is a particular hydrogen ion concentration at which its activity is at a maximum. The work of Soerensen, to which I have already referred, gives a very complete account of this aspect of enzyme action and also of the methods he employed to investigate it. His work should certainly be carefully studied by anyone wishing to take up this part of the subject.
The same remarks as to optimum conditions apply to temperature, although the effect of this is better known. Most tryptic enzymes act best at the body temperature, viz. 98° to 100° F. (37° to 40° C.), and hence puering should be conducted at this temperature. In the case of the hen or pigeon-dung bate the enzymes have not yet been studied, so far as I am aware, so that it is not possible to give an account of them. At the same time it seems highly probable that, as these bates are employed at comparatively low temperatures, the enzyme action is kept back, and therefore the chief action would be a chemical one.
Enzymes are retarded in their action in the first place by their own products, in a very similar manner to bacteria, in the second place the retarding or inhibiting action is brought about by the so-called anti-bodies. Of these the longest known and best studied are those which are produced by toxins, and which neutralize the action of the toxins upon the animal organism (anti-toxins).109 Normal serum also inhibits the action of trypsin and many other enzymes. Another very important group of anti-bodies are the precipitins. If the serum of an animal be injected repeatedly into another animal of different species, a precipitin appears in the serum of the animal treated, which causes a precipitate when added to the serum of the first animal. The special importance of this fact is, that it can be utilized as a method of distinguishing between human blood and that of animals, which is often of importance in medical juris-prudence.110
This fact has been made use of to distinguish dog dung from other matters, with which it has been adulterated.111 A perfectly clear extract of dog dung filtered free from bacteria was injected into a rabbit. The serum obtained from the rabbit was found to contain a precipitin, and on addition of the serum to the dog dung extract a precipitate was produced. When the serum was added to the extract from the dung of another animal no precipitate was formed. When added to the extract of the adulterated dung a very much smaller precipitate was produced than with the extract from dog dung alone.
The coli bacteria in the puer also produce an anti-body—agglutinin.112 If a culture of B. coli be examined under the microscope, the bacteria are seen moving here and there in the liquid, evenly distributed. On the addition of a trace of the serum of an animal which has been previously injected with coli bacteria the bacteria on the slide cease their movements and collect together in clumps. They are said to be agglutinated. This property is used to diagnose bacteria in suspected cases of cholera and some other diseases.
The wonderful discoveries that have been made in this direction constitute one of the most marvellous chapters in the history of science. Serum diagnosis and serum therapeutics are now firmly established as invaluable aids to the physician in his fight against disease and death.
The extent of our present knowledge of the action of enzymes in puering may be summed up as follows. Active enzymes are produced by bacteria growing in the infusion of dung, in addition to digestive enzymes which may be originally present in the dung; the bacterial enzymes are produced more rapidly in a dilute infusion, as employed in the puer wheel, than in the dung itself. The enzymes are of various kinds, proteolytic, peptolytic, lipolytic, etc., but the proteolytic and lipolytic are the most important. These have a solvent action on the fibres of the skin, but little or no action on the hyaline layer, at the concentration usually found in the puer liquor. The fatty matters and soaps in the skin are acted upon by the lipolytic enzymes, and the fats to some extent emulsified, so that they may be easily removed from the skin by scudding or pressing.
It must be clearly understood that enzyme action alone is not sufficient, as has been previously explained, but that the dung enzymes, acting in conjunction with the chemical compounds present, produce the specific puering effect.
There is still much work to be done before the action of the enzymes in dung is fully understood, but from the above short account it will be seen that the part they play in the bate is of great importance.
The reprints of these papers being no longer available, I have thought it best to print them in the present volume exactly as published, with the exception of the paper entitled “Fermentation in the Leather Industry” from which only so much is reproduced as relates to puering and bating.
I. Part of Paper entitled “Fermentation in the Leather Industry.” Read before the Nottingham Section of the Society of Chemical Industry, February 14, 1894.
In the next process, that of “bating,” the bacteria play a more important part, and may be put in our second category, viz. helpful to the tanner, though, from the very nature of the material used, it forms a nidus for various putrefactive ferments, and is therefore dangerous. The materials usually employed are bird dung and dog dung; the former of these is sharp and piercing in its action, while the latter has a more softening action on the skin. In both bates, however, a rapid solution of the gelatinous and albuminous hide substance (coriin) takes place. The hide fibres, however, are not attacked until all the nutrient material in the liquid has been consumed by the bacteria.
Eitner (Der Gerber, xv., 158) states that a sterile bate is without action on the skins, and appears in that article to attribute the entire working of the bate to the action of micro-organisms. He seems to have based his conclusions on experiments with an old bate sterilised with creolin, which he found had no action on the skins. If, however, a fresh bate be made and boiled for half an hour, then allowed to cool to 90°–95° F., it will be found to have a considerable action on the skins, though not so rapid a one as the unboiled bate. The boiling kills all organisms, and fresh ones have not time to develop from spores still remaining before the experiment is at an end.
Lately, the action of unorganised digestive ferments in the dung has been found to be considerable. Thus bating is an exceedingly complex process. The action appears to be threefold:—
| (i) | A purely chemical action of the soluble salts present in the bate on the lime in the skin.113 |
(ii) | An action due to organised ferments. |
(iii) | An action due to unorganised ferments. |
The precise amount of influence each of these actions has on the skin is difficult to assign. The chemical action of the ammonia compounds dissolves the lime remaining in the skin, but the simple removal of the lime is not sufficient, as may be shown by removing it completely with dilute hydrochloric acid or other similar means, and washing perfectly free from acid in distilled water. When tanned, such a skin is hard and brittle.
The organised ferments or bacteria, of which there are many species in the bate, probably act on the skin by secreting soluble ferments, which have the power of dissolving hide fibre. I have isolated in plate cultivations several species which liquefy the gelatin.
If skins are allowed to lie in the bate, zoogloæ of these bacteria collect in the folds and attack the fine “grain,” so that the leather is covered with lines and markings, or “flaked.” Thus the operation is a very critical one, requiring careful watching; under certain atmospheric conditions and at a temperature of 35°–40° C. the skin, if only left a short time too long, will completely melt away. The more skin substance is dissolved in the liquid, the more rapid and pronounced the bacterial action becomes.
So far as is at present known the unorganised ferments in the dung, besides those which are secreted by the bacteria, are mostly digestive ferments114 —pepsin, pancreatin, and trypsin—certain quantities of which pass out of the animal’s body in an unchanged condition. Of these, pepsin only acts in an acid solution, and, although fresh bate liquor is faintly acid to litmus, it quickly becomes neutralised by the lime in the skins, so that the action of this ferment can only be limited in extent. Pancreatin will act in a neutral solution, and has, therefore, a considerable effect on the skin.
Some Dung Analyses.
Hen Dung | Dog Dung | Guano | |||
Per Cent. | Per Cent. | Per Cent. | |||
| Moisture | 60·88 | Ca | 43·049 | Urea | 5·00 |
| Organic matter † | 19·22 | Mg | 0·087 | K2SO4 | 7·90 |
| Phosphates | 4·47 | K | 0·302 | NaCl | 5·00 |
| CaCO3 and CaSO4 | 7·85 | Na | 0·438 | (NH4)3PO4 | 5·50 |
| Alkaline salts | 1·09 | Si | 0·004 | (NH4)2O | 0·60 |
| Silica and sand | 6·69 | PO4 | 3·446 | SiO2 | 2·25 |
──── | CO2 | 7·464 | Ca3(PO4)2 | 9·00 | |
100.00 | Cl | 0·037 | MgNH4PO4 | 15·25 | |
| Fe and loss | 0·008 | Ammon. urate | 15·25 | ||
| Organic matter | 14·152 | Organic matter (17 per cent. N) | 41·73 | ||
| † Containing N = ammonia 0·74 | H2O | 31·013 | Moisture | 6·52 | |
───── | ──── | ||||
100·000 | 100·00 | ||||
W. I. Macadam, J.S.C.I., 1888, 80. Viollet, Diction d’Analyses Chim.
The dog dung actually used in a tannery is from animals fed on a more vegetable diet, the one given having an extremely high percentage of lime, no doubt owing to the animal having eaten bones. A dung as brought from the kennels was found to contain:—
Per Cent. | |
| Mineral matter | 4·679 |
| Organic " | 9·731 |
| Water " | 85·590 |
───── | |
100·000 |
I find by experiments with the purified ferments that their action is very slow compared with the bate itself. Two portions of the same skin were taken: one of them was treated with a 1 per cent. solution of pepsin acidified with 0·2 per cent. of hydrochloric acid, the other in a bate liquor of dog’s dung; both at a temperature of 40° C. (104° F.). At the end of one hour, the skin in the pepsin solution was considerably “fallen,” but that in the manure solution was bated nearly away, i.e. the greater part of it was dissolved.
A 1 per cent. solution of pancreatin (Merck’s) was found to act far more rapidly than pepsin. At 40° C. in a neutral solution the skin fell rapidly, and the action continued even in the cold. In this experiment it was found that in 15 hours the liquid was swarming with minute bacteria. In order to guard against the influence of bacteria a similar experiment, at the suggestion of Mr. H. R. Procter, of the Yorkshire College, Leeds, was tried with the addition to the pancreatic solution of 1·5 per cent. of chloroform; this prevents the development of bacteria, while it does not interfere with the action of the pancreatin. The skin was reduced as before, but in neither case had it the peculiar touch of a “puered” skin, nor were the characteristics of the leather the same.
II. Notes on the Constitution and Mode of Action of the Dung Bate in Leather Manufacture. Read before the Nottingham section of the Society of Chemical Industry, Oct. 26, 1898.
In a paper read before the Society of Chemical Industry (J.S.C.I., 1894, 219), the author gave an outline of the bating process as far as he had then investigated it. Further researches have shown that, although the views then put forward must be somewhat modified, in the main they are correct.
In 1895–96 a series of experiments was undertaken with a view of ascertaining how much of the reducing action of the bate was due to: (1) the chemical constituents; (2) the bacteria; (3) the digestive ferments and enzymes.
The puer used was dog dung obtained freshly every week from hunting kennels. It contained on an average: water, 85 per cent.; organic matter, 10 per cent.; mineral matter, 5 per cent.; about 3 per cent. of the organic is soluble and about 1 per cent. of the mineral matter.
According to Gamgee (Phys. Chem. 2), a dog fed on flesh diet excretes from 27 to 40 grm. of fæces in 24 hours, of which 12·9 grm. are solids. On a bread diet the amount of fæces is much more.
An ultimate analysis gave the following results:—
— | Bread Diet | Meat Diet |
| C | 47·39 | 43·44 |
| H | 6·59 | 6·47 |
| N | 2·92 | 6·50 |
| O | 36·08 | 13·58 |
| Mineral matter | 7·02 | 30·01 |
These figures alone are sufficient to show what variable quantity is the puer employed in a tannery.
The mineral matter is composed as follows:—
| Soluble in water | NaCl and Na2SO4 | 1·37 | 4·00 | ||
| Sodium phosphate | 2·65 | ||||
| Insoluble in water | Earthy phosphates | 80·37 | 94·93 | ||
| Ferric phosphate | 2·09 | ||||
| CaSO4 | 4·52 | ||||
| Silicic acid | 7·94 |
The ammonia compounds combine with the lime remaining in the skin, but the constitution of these compounds was unknown.
The puer was examined for ammonia by Schlösing’s method, 50 c.c. being mixed with 50 c.c. milk of lime placed under an air-tight bell-glass, together with an open vessel containing a measured quantity of standard acid, for 36 hours. No NH3 was given off.115
The mixture of dung and Ca(OH)2 was filtered and a perfectly clear filtrate of a golden-yellow colour obtained. This contained free amines, calcium salts of volatile and non-volatile acids.
It was distilled until the distillate was no longer alkaline, and made up to 500 c.c. 50 c.c. required for neutralisation 5·7 c.c. of (N/100) H2SO4, equivalent to 0·1938 grm. of NH3 per litre of original puer, being the amines set free by Ca(OH)2 in cold solution.
A further 50 c.c. was distilled with NaOH, the amines given off neutralised 8 c.c. of (N/1) HCl, equivalent to 2·72 grm. of NH3 per litre. The neutralised liquid was evaporated to dryness; the residue, consisting of amine hydrochlorates, amounted 0·27 grm. A qualitative test showed both primary and secondary amines to be present.
The action of these amine hydrochlorides was tried upon skin, pieces of limed sheep grain previously washed in water being used. The solution was maintained at 35°C. In the first experiment 0·27 grm. was dissolved in 100 c.c. of water. In two hours the skin was considerably reduced, but had not the touch of a “puered” skin. It tanned well and was a good colour, showing that the lime had been removed, but the leather was not sufficiently soft.
With a strength of 1 grm. per 100 c.c. the action was hastened, the same result being obtained in 1 14 hours, but the coriin of the skin was not dissolved. A comparison piece of the same skin in dung was bated too much.
With a view of comparing the action of analogous bodies, skin was treated in a solution of anilin hydrochloride, 1 grm. per 100 c.c. for 1 14 hours at 35° C. The solution was acid, lime was removed, and the skin felt very similar to that in the previous experiment. Coriin was not dissolved, and the skin did not “come down.”
Of the mineral constituents mentioned above, only the chlorides116 have a reducing action on skin, the other compounds being inert, e.g. silicates, or merely supply food for the bacteria.
For a general outline of the decompositions taking place in dung, see Dr. Herfeld’s paper (J.S.C.I., May 1895). We need only consider the action of the chief organic compounds, as the amount of chlorine is too small (in one case 0·053 per cent. on the dry dung) to be of importance. The principal compounds are:—
Organic Acids.—Formic, acetic, butyric, valerianic, lactic, malic, tartaric, citric, and glyceric.
Amido Compounds.—Leucin, tyrosin, glutamic acid, glutamine, asparagin, glycocol.
Bases, consisting of amines, skatol, and indol, and varying amounts of ammonia, according to the age of the dung.
The organic acids exist partly as salts of the alkaline earths, partly combined with amines. A distillation of “puer,” after acidifying with H2SO4, gave 2·2 grm. per litre of volatile acids reckoned as acetic. No HCl came over. On decomposing the sodium salts of the acids with H2SO4, the smell of butyric acid predominated. I have also shown the presence of lactic acid in the dung, but have been unable to determine the amount.
Action of Amido Compounds.—A mixture of glycocol and leucin117 was prepared by boiling 8 grm. of gelatin in 400 c.c. of water, acidified with 1 c.c. of HCl for two hours with inverted condenser. The acid was then neutralised with ammonia. The solution had a very considerable reducing action on skin at 35° C. in 90 minutes, but not sufficient for practical purposes. It appears to dissolve a little coriin in a similar way to dilute acids. It was found on further investigation that the action was largely due to the NH4Cl present.
Action of Dilute Acids.—Although the bate in practice is almost always alkaline, yet a fresh bate is acid, and it may be as well, before proceeding, to consider briefly the action of weak and dilute acids on skin.
The fibres of the skin have only a limited capacity for holding acids, and soon begin to swell abnormally and partially dissolve. Reimer118 has found that the material may be reprecipitated by lime water. It forms a fibrous mass, which has not the sticky feel of gelatin, but is at once converted into that body by boiling.
The author has examined the action of dilute H2SO4 on sheep skins, and this may be considered typical of the action of acids on skin generally.
When puered and drenched sheep skin is treated for 20 minutes with a very dilute solution of H2SO4 ( 1280), it swells up considerably and becomes soft and semi-transparent, The fibres of the skin, which under normal conditions have a diameter of 4µ–6µ become 20µ or more; on staining with picro-carmin there are seen to be certain fibres which are unaffected or only slightly affected by the acid. These are the elastic fibres119 and the capillary blood vessels.
On filtering a solution of the above strength in which skin in excess has been treated for 20 minutes, and evaporating 100 c.c. to dryness in a platinum dish, a residue of 0·4992 grm. was obtained. A second experiment gave 0·496 grm. of which 61 per cent. was organic matter.
By Kjeldahling another portion of the residue, the N in 0·12 grm. amounted to 0·0122 grm. equivalent to 0·073 grm. of skin substance; i.e. 60 per cent. of the soluble matter is skin. It is evident that the matter in solution is a compound of skin substance and H2SO4 together with a little soluble mineral matter contained in the skin. The amount of substance dissolved depends upon the condition and previous treatment of the skin.
In the presence of NaCl, which prevents the swelling of the fibres, puered sheep grains take up 7·0 c.c. of (N/1) H2SO4 from a solution containing 17 c.c. of (N/1) H2SO4 (0·833 grm. H2SO4) and 7·8 grm. of NaCl per 100 c.c. About 300 grm. wet skin per litre were used. The skin substance dissolved by the acid in the time named does not appear to be more than that dissolved by the weak acids of a drench, although an equivalent weight of H2SO4 has a much more powerful swelling action on skin than these acids.
A piece of puered and drenched sheep skin was carefully washed in distilled water and then dried in vacuo until it ceased to lose weight; the absolute dry weight was 4·2670 grm. The skin was soaked down in distilled water and again drenched for 30 hours in a clean bran drench, developing approximately 0·8 grm. lactic acid, and 0·5 grm. acetic acid per litre (see this Journal, May 1893). It was again washed and dried in vacuo. It weighed 4·12 grm., a loss of 0·14 grm. = 3·44 per cent.
The action, therefore, of the acids is a reducing one, inasmuch as skin is dissolved, but as the fibres hold a certain amount of acid, the skin appears to be plumped. Where the acid is a mineral one the skin tans plump, but with a brittle and inelastic fibre, weak organic acids make a plump, soft leather with a moderately elastic fibre, giving to the skin a somewhat india-rubber feel. We thus see that weak acids alone do not produce a similar result to a dung bate, which causes the skin to fall and produces a flat soft leather with a stretch in it, which will not spring back.
I have also tried the action of the following Na and NH3 salts upon skin:—
Sodium lactate, C3H5O3Na, prepared by neutralising 2 grm. lactic acid with sodium carbonate in 1000 c.c. water. The solution was used under the same conditions as the other bating experiments, viz. at 37° C. the reducing action was nil, but on prolonging the digestion, the medium was found to be very suitable for the development of putrefactive organisms, and, as a consequence, the skin was attacked and partly peptonised.
Ammonium lactate, prepared in a similar way, had an almost identical action on the skin, but was more effective in removing the last traces of lime.
Ammonium butyrate, prepared as above, removed lime, but instead of a reducing action it appeared to plump the skin slightly.
Action of Bile.
Amongst other compounds in the dung which might have some action on skin were the bile salts and bile colouring matters. (For a full description of these, see Gamgee, Phys. Chem. ii.)
Experiments were made with ox bile, 25 c.c. being diluted with 250 c.c. of water. Pieces of sheep grain previously washed in water were digested in this solution at 37° C. for 1–4 hours. The bile had no reducing action on the skin and in fact hardened it, at the same time staining it a dirty yellowish brown, the colour differing from that of the colouring matter of dung (hydrobilirubin). Even after continued standing there was little or no development of bacteria, the bile appearing to have an antiseptic action.
Action of Bacteria.
Having ascertained the action of the chief chemical constituents of the dung, that of the bacteria was next examined. In the first experiment, eliminating all but bacterial action, a tube of nutrient gelatin was inoculated from an active bate. In two days the gelatin had liquefied along the needle track.
The culture was now transferred to the following solution:—
| Gelatin | 4 | grm. |
| Dextrose | 4 | " |
| K2HPO4 | 1 | ·0 " |
| MgSO4 | 0 | ·2 " |
| NaCl | 0 | ·4 " |
| Water | 2000 c.c. | |
The growth in this was fairly rapid and the smell very slight at the end of a week. A piece of washed sheep grain worked in this culture at 37° C. for four hours was considerably reduced, the grain was not attacked, the action differing in this respect from a prolonged puering. This mixed culture was grown in various other nutrient media, gelatin, gelatin and mineral salts, broth made from sheep and calf fleshings, etc., with practically the same results. In no case did the bating action equal that of dung, and was insufficient for practical purposes.
A cursory bacteriological examination of dung will show that the number of species of bacteria it contains is very large, but on making a plate cultivation from a sufficiently dilute infusion of fresh dog dung, most of the colonies developing appear to belong to four or five species. The following is a list of all the organisms which, so far as my knowledge goes, have been isolated from different kinds of dung:—
List of Known Bacteria in Dung.
| 1. | Bacillus fluorescens putridus | These organisms have been formerly described as Bacterium termo by various authors. | |
| 2. | " " liquefaciens | ||
| 3. | Proteus vulgaris. | ||
| 4. | " mirabilis. | ||
| 5. | Bacillus subtilis. | J. T. Wood, J.S.C.I., 1890, 28. | |
| 6. | Micrococcus fulvus. | ||
| 7. | Bacterium ureæ. (Cohn.) | ||
| 8. | Bacillus saprogenes, I., II., III. Vide Herfeld, J.S.C.I., May 1895. | ||
| 9. | Bacillus butyricus. | ||
| 10. | B. putrificus coli. | ||
| 11. | B. pyocyaneus. | ||
| 12. | B. janthinus. | ||
| 13. | B. coprogenes fœtidus. | ||
| 14. | B. pyogenes fœtidus. | ||
| 15. | B. prodigiosus. | ||
| 16. | B. zenkeri. | ||
| 17. | B. magnus. | ||
| 18. | B. spinosus. | ||
| 19. | B. liquefaciens. | ||
| 20. | B. coli. | ||
| 21. | B. duclauxii = B. ureæ. | ||
| 22. | Micrococcus ureæ. | ||
| 23. | " " liquefaciens. | ||
| 24. | Proteus sulphureus. | ||
| 25. | Bacterium sulphureum. | ||
| 26. | Bacillus amylobacter. | ||
| 27. | Clostridium butyricum. | ||
| 28. | B. acidi paralactici. | ||
| 29. | Bacillus I. | Isolated from horse manure by S. A. Severin, Centr.-Bl. f. Bakt. ii. [1], 97. | |
| 30. | " II. | ||
| 31. | " III. | ||
| 32. | Spirillum serpens. | Isolated by Kutscher, Zeit. f. Hyg. xx. 45. | |
| 33. | " tenue. | ||
| 34. | " undula. | ||
| 35. | " volutans. | ||
| 36. | Vibrio I. | ||
| 37. | " II. | ||
| 38. | " III. | ||
| 39. | Spirillum from pig dung. Smith. Centr.-Bl. f. Bakt. xvi. [1], 124. | ||
| 40. | Streptococcus liquefaciens coli. Gamgee. Phys. Chem. ii. | ||
| 41. | Bacillus mycoides.120 | ||
There are besides many unnamed species of bacteria in dung, among which are the two following isolated by me from puer—
(a) Plate Cultures.—Small yellowish colonies, slightly fluorescent, liquefy the gelatin slowly. Rods resembling B. subtilis when at rest, but moving with a rapid undulatory motion. Cultures in nutrient gelatin have a considerable reducing action on skin.
(b) Plate Cultures.—Bluish colonies liquefying gelatin rapidly, micro-bacteria in pairs. Not so marked an action on skin as (a). The organism resembles Proteus vulgaris, but swarming islets not observed.
Of the species mentioned above, a pure culture of B. subtilis in meat broth, maintained for one week at 35° C., had no reducing action on skin. B. fluorescens liquefaciens had a moderate reducing action.
The species developing on the plates vary according to the age of the dung. In horse manure, according to Severin,121 bacilli predominate in the early stages. At the end of two to three weeks micro-bacteria, cocci and diplococci make their appearance, while the bacillar form becomes scarce. In three months cocci predominate, forming zoogloæ; strepto- and staphylo-cocci and spirilli were seldom found. Yeasts and sarcina were never observed. From observations extending over several years it is evident that a similar cycle occurs in dog dung. When quite fresh it contains comparatively few bacteria; then certain groups of species take possession, causing decompositions in the dung; these in their turn give place to other species, which decompose the products formed by the first, so that no one species produces or can produce the complex chemical and physiological changes which take place, or the bodies necessary for the bating of skin, as has been supposed by some observers.
None of the experiments, either with mixed or pure cultures of bacteria, had a perfect reducing action on skin, although the action was considerably greater than that of the chemical solutions. It was found, however, that by adding a small quantity of the amines above mentioned to the bacterial cultures, the reducing action became nearly as rapid and effective as with dung itself; and it was, therefore, clear that the principal bating effect was due to a combination of two things, viz. an active growth of bacteria in the presence of amine compounds.
It was now necessary to ascertain the mode of action of the bacteria on the skins.
I have previously shown122 the action of digestive ferments or enzymes to be notable in the case of pancreatin. A careful study of the behaviour of the digestive enzymes in the animal body shows, however, pretty conclusively, that they are all destroyed before the fæcal matter is discharged,123 and, therefore, the enzymes in the dung are not from this source.
An attempt was made to filter the dung diluted with water through a Berkefeld filter, and thus obtain a clear filtrate containing all the enzymes and free from bacteria; it was found impossible to filter the liquid in this way.
The method of Claudio Fermi124 was also tried, but a clear filtrate could not be obtained.
The author has succeeded in obtaining the enzymes best in the following manner:—About 150 c.c. of the above-mentioned “puer” was well mixed with an equal quantity of glycerin and allowed to stand for seven days; it was then capable of being filtered on a filter pump, although very slowly, and yielded a clear filtrate of a deep golden brown colour; the filtrate was poured in a thin stream into a tall vessel containing about 1500 c.c. of 98 per cent. alcohol. A copious flocculent precipitate of the albuminous matters and enzymes was thrown down, the alcohol was filtered off and the precipitate washed on the filter with absolute alcohol, and dried over sulphuric acid. The resulting amorphous body was of light brown colour, and became darker when exposed to the air. The amount obtained from 150 c.c. of dung was 0·55 grm. (3·66 grm. per litre). The body consists of a mixture125 of all the enzymes existing in the dung along with other albuminous bodies. It has a very slight diastatic action on starch. 0·5 grm. in 100 c.c. of water at 35° C. had a very considerable reducing action on skin. A further experiment with 0·5 grm. of amine hydrochlorides, 0·5 grm. of enzymes and 100 c.c. of water, at 35° C. brought down a piece of limed sheep grain in 30 minutes exactly like puer; the reaction of the solution at the commencement of the experiment was faintly alkaline; at the end of the experiment it was considerably alkaline.
It was noted that the fæcal odour of the glycerin solution disappeared on standing for two or three weeks, and the solution smelt strongly of ethyl butyrate, the enzymes being still present.
These enzymes were prepared from dung, and it now remained to prepare them in the laboratory by the action of bacteria alone. For this purpose 200 c.c. of a mixed culture of dung bacteria in the solution above described, seven days old, was mixed with 200 c.c. of dilute alcohol (alcohol 65, water 100) and well shaken. Gelatin and albuminoid bodies are by this means precipitated. The liquid was filtered and poured into eight times its volume of 98 per cent. alcohol. The precipitate which came down was washed with absolute alcohol and dried in the usual way. The enzymes thus obtained were redissolved in water and the former experiment with skin repeated with this solution. The same effect was produced, showing conclusively that it is the enzymes produced by the bacteria, acting in conjunction with the amines, which bate the skin. It would seem that the special action of the enzymes is aided by the presence of amine compounds, in addition to the chemical action which these latter have upon the skin. The action is shown to be interdependent, i.e., bacterial action alone is insufficient, and chemical action alone is insufficient, the true bating action being a combination of the two.
In conclusion the author has pleasure in acknowledging the valuable assistance of Mr. H. S. Shrewsbury in carrying out some of the experiments.
III. Further Notes on the Action of the Dung Bate.126
The present notes are a continuation of those published in 1898. It has been impossible for me to follow up the subject except in a desultory manner, but I think it desirable to put further results, however meagre, before you with the object of throwing more light on the complex process of bating hides and skins.
Since the previous notes were written, Dr. Th. Körner127 has called attention to the physics of the process, a matter which I had not previously mentioned. He explains part of the action by differences of osmotic pressure. The ammonium salts and salts of organic bases contained in the dung, possess a smaller osmotic pressure than the solution of calcium hydrate contained in the skins. The ammonium ion endeavours to unite with the hydroxyl ion of the calcium hydrate to form undissociated ammonium hydroxide. In consequence of the osmotic pressure and the electrostatic attraction of the oppositely charged ions, the calcium hydrate solution is withdrawn from the skin and the latter falls.
Experiments with Bacteria on a Large Scale.
I have shown128 that simple cultures of bacteria in certain nutrient media, in the absence of amines and other bodies, have an imperfect bating action on skin. This was the case, whether the cultures were pure or mixed, but the mixed cultures have a better action than the pure cultures of any of the organisms I have hitherto used. Bacteriological processes in nature are usually carried on by a mixture of species of bacteria; still there are fermentations like the spontaneous souring of milk, the formation of nitrites from nitrates, and the ammoniacal fermentation of urea, which may be considered as natural pure cultures. These are examples of what I may call the selective influence of the nutrient medium. The nutrient medium used in my former experiments was a solution of gelatin and mineral salts, such as is used in general bacteriological work, modified in various ways. On careful consideration, this did not appear to be a medium at all corresponding to the natural bate. The albuminoids in the process of digestion are peptonised, and dung contains only those bodies which the animal is incapable of assimilating. It is from these bodies that the necessary enzymes are produced by bacteria. Of the numerous trials with pure cultures of different organisms from dung, I found that most of those having the property of secreting bating enzymes, were non-liquefying bacteria. Popp and Becker have also pointed out that peptonising bacteria do not exert a favourable reducing action on skin. It is impracticable to grow these non-peptonising organisms on a large scale in a solid medium, and it occurred to me that gelatin, previously peptonised by chemical means, would be a favourable medium. Experiments with hydrochloric and sulphuric acids gave hopeful results. The acid was neutralised with ammonia after the chemical action was complete, and the bacteria cultivated in this solution.
A better result was obtained by digesting 10 grm. gelatin, 5 grm. lactic acid (anhydrous by calculation), 100 c.c. of water, in a closed vessel on the water bath for three hours. A slight black precipitate of melanoidic acid (humic acid) is formed. The liquid is of a clear brown colour, and contains a large number of nitrogenous bodies resulting from the breaking down of the gelatin molecule. It has been impossible for me to ascertain its exact composition, but I have partially examined it with the following results:—
1. A small quantity of the liquid poured into absolute alcohol, gives a white precipitate which re-dissolves on the addition of about 30 per cent. water. This shows absence of gelatin.
2. On saturating with ammonium sulphate, a brownish precipitate is thrown down which is perfectly soluble in cold water. It consists of gelatose lactates which are evidently formed in an analogous manner, to the gelatose hydrochlorides prepared by C. Paal.129 The gelatoses correspond to the protoses formed during the digestion of albumin. In one sample 1·3 grm. gelatose lactates per 100 c.c. was found.
3. The filtrate from (2) was dialysed against running water for 15 hours to get rid of the ammonium sulphate. The resulting gelatin-peptones, or gelatones, amounted to 9·1 grm. per 100 c.c. In other words, the heating under pressure had transformed 87 per cent. of the original gelatin into true peptones.
It may be presumed that the other and simpler nitrogenous bodies formed, are the same as those produced by the treatment of gelatin solutions by dilute mineral acids, and that the bases combine with the free lactic acid. An excellent list of these and a very interesting account of the protamines and hexones which go to form the complex albumin molecule is given in a paper by Dr. A. Kossel.130
The solution of gelatone lactates and free lactic acid prepared as above, was neutralised with sodium carbonate and diluted to 1000 c.c.; it was found to be a very good medium for the growth of the bating organisms after the addition of a small quantity of potassium phosphate.
In looking round for suitable means of testing the action of both pure and mixed cultures of bacteria on a practical scale, it occurred to me that the Carlsberg vessels, as used by Hansen for the pure cultivation of yeast,131 would answer for the cultivation of bacteria also. I was unable to obtain any information on the subject, and was surprised to find that the apparatus is very little used in England. I procured two of these vessels from Jensen of Copenhagen, and found they answered very well. The bacteria were transferred from the test tube in which the original inoculation had been made to a Pasteur flask of 250 c.c. capacity containing the nutrient solution; when the growth in this flask was sufficiently vigorous the Carlsberg vessel was inoculated from it in the manner described by Hansen, and with all the usual precautions.
After three days at a temperature of 37° C. the whole of the 10 litres was used for inoculating 100 litres of the nutrient medium above mentioned contained in a clean barrel standing in a room, the temperature of which was maintained at 37° C. By using this comparatively large volume of pure culture for pitching, if I may be allowed to use a familiar brewing term, the large culture was kept practically pure, although it could not be supplied with germ-free air, as in the case of the Carlsberg vessel.
Using a nutrient medium containing 1 per cent. original gelatin, the maximum bating effect was obtained with the majority of organisms in three days.
In my previous notes I showed132 that a mixture of digestive enzymes and amine hydrochlorates was effective in bating skin. The liquid in this case was free from bacteria. It was found, however, that the action of the bate was hastened when an active growth of bacteria was going on at the same time in the liquid, even though the quantity of enzyme present was smaller. This is what one might infer from a careful study of the process. It appears better for small quantities of enzymes to be produced in the liquid as required, than to use larger quantities of stale enzymes.
Mr. Loxley Meggitt was kind enough to concentrate for me about 170 litres of a culture of bating organisms. The concentration was conducted in vacuo at a temperature below 50° C., at which the enzymes were certainly not injured, and it was found that when this concentrated culture was diluted to the same strength as the original, the action was considerably diminished. This diminution of activity was due in part to a loss of volatile products in the evaporation, but more especially to the absence of an active fermentation going on in the liquid.
The nutrient medium above described, besides acting in a selective manner on a mixture of bacteria, and providing them with food stuff, contains amido compounds and other bodies, which are eminently favourable to the proteolytic action of the enzymes, and therefore act in the same way as the amine hydrochlorates which I used in former experiments.
The action of the following bacteria grown in this medium has been tried on skin. The pure cultures are lettered in continuation of previous descriptions (see previous paper, p. 162).
1. Bacillus c, isolated from pigeon dung bate. Small, round, bluish-white colonies, standing up slightly above the surface of the gelatin, but not spreading out on the surface, non-liquefying pairs of micro-bacteria. This was the principal organism found in a pigeon dung bate as used for the bating of E.I. kips. Grown in nutrient broth or in the special medium it had no reducing effect on skin.
2. Bacillus pyocyaneus from blue skin, i.e. skin in an early stage of putrefaction. No action.
3. Mixed culture from fresh pigeon dung, collected in a sterile vessel. This contained very few bacteria capable of developing in the special medium. The action on skin was no more than that due to the chemical compounds present.
4. Mixed culture from fresh dog dung. This contained remarkably few species of bacteria which came to a good development in the special medium. The principal organisms were a jointed bacillus with extremely rapid vibrionic movement, and a small bacterium in pairs; the action on skin was perceptible but slight.
5. Mixed culture from dog dung one month old, as used in the bate. The action of this in a certain medium has already been described (see p. 160). In the special medium it was distinctly better and almost equal to that of the dung bate.
6. Mixed culture from fresh fæces gave a similar result to (3). A microscopic examination showed pairs of small bacteria and micrococci, but no bacillar forms.
7. Mixed culture from fresh horse dung had a moderate but distinct action on skin, about equal to (4).
8. Bacillus d, isolated from wool infusion. Very slight action.
9. Bacillus e, isolated from wool infusion. Similar result to (8).
10. Mixed culture of bacilli d and e only. Very powerful bating action, skin bated more rapidly than with dung.
These experiments tend to confirm the conclusion I previously arrived at, viz. that no single species of bacteria produces the complex chemical and physiological changes which take place in the dung, and which result in the production of the bodies necessary for the proper bating of skin. It is a well-known fact that dog dung requires keeping for at least a month before it gives the best result. During this time it undergoes a kind of fermentation and continues to improve up to two months, after which it deteriorates. The rate of fermentation depends upon the season.
It is evident from these facts that the bacteria present in the dung when it leaves the animal’s body do not produce the required enzymes and chemical compounds, and that these are produced by bacteria which obtain access from the air. It is also evident that the production of these enzymes depends upon the composition of the nutrient medium, since this exerts a selective influence on the species of bacteria obtaining access to it. In the spontaneous souring of milk numerous bacteria in the air have free access to the milk, yet the lactic ferment is generally so pure that it may be and is used as a pure culture on a large scale in the manufacture of lactic acid. Dog dung is a favourable medium for the bacteria secreting the bating enzymes; but if dextrose or other carbohydrate be added to fresh dung an acid fermentation is set up which effectually prevents the development of the bating organisms.
Reasoning along these lines I was led to try some cultures of air bacteria in my special medium, and I found that a good source of such organisms likely to have a useful effect was a sweating stove as used for the depilation of skins. The hair roots are loosened by bacterial action; the wool, when it slips, brings away with it the epidermis. The root portions of the wool were cut off and digested in water at 35° C., the liquid strained off and used for making plate cultures in the usual way. In making the attenuations for the plate cultures, the fourth was found to be practically a pure culture of the organism I have called bacillus d, or the sweating bacillus. It forms large whitish colonies, spreading on the surface, with irregular contour. The bacilli are very small, mostly in pairs, but sometimes joining together in thread-like forms. (Fig. 21.)
Grown in the special medium it had little or no action on skin. At the same time I found that a culture made from the original liquid, i.e., a mixed culture of the sweating organisms, had an exceedingly powerful bating action; indeed, the skin was bated more rapidly than with dung.
All the experiments carried out so far tend to prove that mixed cultures of suitable bacteria possessed the required action, whereas pure cultures do not. A further examination of the infusion of wool roots showed that the bacteria contained in it, consisted practically of two species only. The first of these I have already described, the second one, bacillus e (Fig. 22), forms small brownish-yellow boat-shaped colonies on gelatin plates, very similar to one of the dung bacteria. They consist of plump cells, two or three times the size of bacillus d, united in pairs and short chains and surrounded by a capsule; the cells appear to vary considerably in size. Cultivated in the same way as the others it has very little action on skin. It is evident from these facts that the growth of the bacteria is a symbiotic one; separately they exert little or no action, whereas used together the action is most remarkable.
The author wishes to express his indebtedness to Mr. J. Golding, F.I.C., of University College, Nottingham, who made the first pure cultures of these “wool” bacteria.
The Influence of Solid Matter in the Bate.
In making comparative tests of my artificial bates and dog dung, I usually took the latter from the paddles in which it was diluted ready for use, and maintained both solutions at exactly the same temperature. Parts of the same skin were then digested in each solution, and the results noted after 1 12 hours. In order to make them more strictly comparable I filtered the dung bate, so as to employ the matter actually in solution when the bate was made up. It was found that the filtered bate had far less action than when it was used in an unfiltered condition. That this is not wholly due to the passing into solution of some of the solid matter during the bating was shown by adding an inert solid, viz., kaolin, to the filtered bate, when the action was greatly hastened. On adding kaolin to the artificial bate and keeping the liquid agitated, the same result was obtained. It seems that the finely-divided solid matter acts as a carrier for the enzymes, perhaps by a kind of mass action, each particle offering a surface vastly greater than the molecules of dissolved solids. In the case of “puer,” the organic insoluble matter is gradually brought into solution by bacterial action as the bating proceeds, though I do not believe this occurs to any great extent.
The following quantities of soluble and insoluble matter were found in puer wheels in actual practice per 100 c.c.:—
— | 1 | 2 | 3 | 4 |
| Total solids | 10·20 | 8·63 | 8·64 | 3·26 |
| Soluble | 6·00 | 4·57 | 6·19 | 2·14 |
| Insoluble | 4·20 | 4·06 | 2·45 | 1·12 |
1. Wheel in constant use for a week.
2. Wheel freshly made up.
3. The same wheel after one lot of skins.
4. From pigeon dung bate pit.
On comparing 2 and 3 it will be seen that apparently 1·61 grm. of the insoluble matter has passed into solution. This is not entirely the case, for part of the additional soluble matter is lime and hide substance from the skins, while part of the insoluble at the same time adheres to the skins and is not estimated.
In conclusion, it will be seen that it is now quite possible to produce an artificial dung bate, or rather a bate having the essential properties of a dung bate, by producing the required enzymes by fermentation and then adding to them the amine compounds. Such a bate I am able to show you here. The chief difficulty in its practical application in the tannery is the question of cost. If this can be overcome, and efforts are being made to overcome it, there is no doubt that the tanners of the next generation will be supplied with a bate which will do the same work as the dung bate, and which will have a definite and uniform composition and properties, and a regular and certain action. The tannery will become cleaner and healthier, and what is perhaps of equal importance, the sewage effluents will be comparatively harmless.
As may be supposed, inventors have from very early times turned their attention to the substitution of some other material in place of the unappetizing one so long in use. But as I pointed out in a short article entitled “Rationale of Bating”133 many of them looked upon bating and puering as a process for removing lime from the skins, and nothing more; whereas we have seen that it is a very complicated process in which, not only is lime removed more or less effectively, but the skin fibres are acted upon, and portions of them rendered soluble, and the skin thus got into the necessary condition.
This misconception of the object of bating or puering prevented the success of most inventions of artificial bates up to quite recent times. Procter states that the use of ammonium chloride as a bate was patented by Zollikoffer in 1838. Such agents as this, and various acids, which have been proposed are merely chemical deliming agents, and, as such, we do not propose to discuss their use as artificial bates. Procter134 gives an excellent account of these processes, including one of his own, in which ammonium chloride is used in conjunction with metabisulphite of soda, and for successive packs sufficient sulphuric acid is added to neutralize the ammonia formed.
The same author135 also mentions an American invention for bating by the use of old lime liquors neutralized with sulphuric acid. This method is much more scientific than would at first sight appear, and more nearly approximates to the conditions of the dung bate than any of the early inventions. Old lime liquors contain much ammonia and weak organic acids, such as caproic, amido-caproic (leucin) and tyrosin. On adding sulphuric acid, the lime forms an inert sulphate, and the sulphate of ammonia and the weak organic acids which remain dissolved are just what are required in a chemical bate. The author has used this bate with success on certain classes of goods.
Tiffany’s bate, which is used in America, is composed of glucose and glue, and bates of this composition are also in use in England. As an example, 10 lb. glucose and 12 lb. of glue dissolved in 1000 gallons water, at a temperature of 75° F., and allowed to ferment for a few hours, may be used for the bating of bellies, and if used in a paddle or latticed drum will bring them “down” very rapidly.
Davis136 gives a list of twenty-nine patents for bating issued by the U.S.A. Government from 1790 to 1883 inclusive. I do not know whether any of these are in use at the present day.
Erodin.—Since the publication of the authors researches on the constitution and mode of action of the dung bate, renewed attention has been given to the construction of bates, or puers, having an identical action. One of these has been worked out by Professor Dr. H. Becker, of Frankfort o/M, in conjunction with the author under the name of Erodin (Latin, erodere, the equivalent of the German beizen—to bate).
As this was the first artificial bate in which a pure culture of bacteria has been applied to the treatment of skins, I propose to describe it in some detail.
Erodin137 consists essentially of a nutrient material in the form of powder and of a pure culture of B. erodiens which is added to the powder mixed in a definite quantity of warm water (40° C.). On the average about 10 grm. of this material is sufficient to bate one kilo of wet prepared skin.
In practice it is found that this material acts far more energetically on the skins when living bacteria are present in the liquid; we suppose that they penetrate the skin and form their products in the intercellular spaces. It was found that, in the evaporation of the cultures to dryness, the bacteria were mostly killed or so far reduced in activity, that on preparing the bate after a certain lapse of time it was liable to become infected by foreign organisms.
In order to ensure uniform results a pure culture of the special bating bacteria is supplied with every 5 kilos of the erodin powder. The weighed amount of erodin is placed in a perfectly clean mashing-tub, with 50 times the weight of water at a temperature of 40° C. a pure culture of Bacillus erodiens is added to the mixture, and the whole covered with a clean cover and allowed to stand for two or three days. A little steam is admitted morning and evening so as to maintain the temperature at about 40° C., or some other suitable device for heating is applied. Several tubs may be kept in a small stove heated to the right temperature by steam pipes. In this way the bating bacteria develop rapidly, and if the proper precautions as to cleanliness and temperature have been observed, there is no fear of any adventitious germs getting the upper hand. As soon as a vigorous growth is attained, usually on the third day, the bate is ready for use.
Erodin is in use on a fairly large scale, and, as is usually the case, difficulties have arisen on a manufacturing scale which do not occur in the laboratory. In the practical application of the material the mashing-tub may be 3 ft. 9 in. in diameter and 3 ft. 3 in. in height and of a capacity of 200 galls. Steam is admitted directly by means of a copper pipe fitted with a boiling jet.
The goods are bated in the ordinary bating paddle or puer wheel (Fig. 3, p. 13).
During the year 1901 about 9000 dozens of sheep “grains” were puered with Erodin at Trent Bridge Works (Messrs. Turney Brothers, Ltd.) Nottingham, and considerable experience gained in its use.
The mode of operation was as follows:—On Fridays 30 kilos of Erodin powder was mashed in a wooden vessel in 1600 litres water at 40° C. and inoculated with a sufficient quantity of a pure culture of B. erodiens. The temperature was kept at 35°–40° C. by admission of a little steam first thing in the morning, again at noon, and in the evening, and the liquid so prepared was ready for use on Monday. One wheel or paddle was kept going for the work. To start the operation, 200 litres of stock liquor was put into the wheel, and sufficient water added for the goods to turn freely. The skins previously washed, as described in Chapter I., were paddled until sufficiently “fallen” or “down.”
For the following lots, the liquor was strengthened by the addition of 100 to 140 litres of the stock liquor, the wheel being run off and cleaned out once a week. The general result showed that “high-limed” skins and previously salted skins were more refractory to this bate than to dung.
As in the English system of collecting pelts there are all stages of liming, skins containing from 1.5 to 9 per cent. CaO on the dry skin, it is not surprising that considerable difficulty was experienced in keeping the puering uniform. Skins containing over 3 per cent. CaO on the dry skin, seem to require a real putrefactive action in order to “bring them down” sufficiently, or it may be that their alkalinity is too great to allow the bating enzymes to act properly. It may here be said that if the fibres of skin are actually damaged by excessive action of lime or any other chemical, no subsequent treatment is of avail to remedy the defects.
Shearling pelts were excellent when puered with erodin, a further proof that the alkalinity is of importance, since these pelts do not require so much lime as “old fells” to bring them into proper condition for splitting.
For sheep and calf skins treated from the market condition and consequently containing a uniform amount of lime—2–3 per cent.—the process is quite reliable, and is in practical use on a large scale. Goat skins have been successfully bated with erodin by treating them in a concentrated liquor in a drum,138 instead of a paddle, the mechanical effect of the drumming enabling the bate to penetrate the hard and compact skin, while the more concentrated solution of the enzymes attacks the so-called “beard,” and renders the skins as supple as sheep skins bated in a paddle.
It may be of interest to give here some of the results of the early trials of erodin in a tabular form, due to the late Franz Kathreiner. They are a further proof of the thoroughness with which he carried out all his work.
Leather, N.F. and Kalochrom. Date of Trial, Nov. 22, 1898.
Remarks.—22 skins of E 47 after 3 hours running had 250 c.c. 70 per cent. lactic acid; they were marked with a hole in the tail; were more fallen than the others.
Judgment after Scudding.—All lots well down; E 47 whitest colour; all smooth grain, E 47 and B 3 smoothest. The 3 goat were not low enough.
It was observed both in laboratory experiments139 and in practical work that the presence of solid matter in the puering liquor had a considerable influence on the speed at which the goods fell. The clearer the liquor the slower the action. The explanation of this fact is not quite clear. Possibly the finely divided solid matter acts as a carrier for the enzymes, each particle offering a very extended surface, or it may be that the action of the solid matter is mechanical, like that observed in bran drenching.
The goods puered with erodin, whether split or unsplit skins, should be scudded on the grain. Sheep splits are scudded with a machine of Sir John Turney’s type. (Fig. 6, p. 17.) Roans and goat may be scudded with any of the spiral knife machines (Turner, Mœnus, etc.) in general use.
The following figures, p. 187, giving the reactions of an erodin bate in c.c. N/5 alkali or acid required to neutralize 50 c.c. filtered bate, are of interest as being taken from actual work. They were kindly communicated to me by Herr Karl Schorlemmer, chemist to Messrs. Doerr and Reinhart, of Worms.
Erodin, tested by the electrometric method, was found to have the following hydrogen ion concentrations:—Paddle before goods, π = 0·672 volt, equivalent to a hydrogen ion concentration p+H 6·6. The same liquor after goods, π = 0·710, equivalent to p+H 7·3. Compare puer liquors, p. 79.
Bate | Before Goods | After Goods | ||
1. | Old | 0·05 alkali | 0·4 acid | |
2. | Fresh | 0·15 | " | neutral |
3. | Old | 0·05 | " | 0·55 acid |
4. | Fresh | 0·2 | " | 0·05 " |
5. | Old | 0·1 | " | 0·1 " |
6. | Fresh | 0·45 | " | 0·15 alk. |
7. | Old | 0·15 | " | 0·3 acid |
8. | Fresh | 0·5 | " | 0·1 alk. |
9. | Old | 0·1 | " | 0·3 acid |
10. | Fresh | 0·3 | " | neutral |
11. | Old | 0·1 | " | 0·7 acid |
12. | Fresh | 0·2 | " | 0·05 alk. |
13. | Old. | 0·1 | " | 0·25 acid |
14. | Fresh | 0·4 | " | 0·15 alk. |
15. | Old. | 0·05 | " | 0·3 acid |
16. | Fresh | 0·45 | " | 0·1 alk. |
| Mean of old | 0·087 | " | 0·31 acid | |
| Mean of fresh | 0·33 | " | 0·051 alk. | |
Oropon.—An interesting application of the theory of the bating process which was put forward by me140 is that of Dr. Otto Röhm141 in 1908, in which, instead of a bacterial culture, he employs an infusion of pancreas in conjunction with ammonium salts. This bate has been put upon the market in the form of a powder under the name of Oropon.
The approximate composition is:—
Per cent. | |||
| Ammonium chloride | 65 | ||
| Wood fibre | 31 | ||
| Dry pancreas | about | 3 | ·5 |
The nitrogen unaccounted for by ammonium chloride is 0·32 per cent.
Its chief advantage is the simplicity of application, no previous fermentation being required. About 5 oz. to 9 oz. are required per 100 lb. wet pelt, and it is recommended to soften the water by adding a certain quantity of clear lime water to it previous to making up the puer.
Its disadvantages are, that unless a previous bath is used, the quantity of ammonia given off is detrimental to a proper bating action, so that for good working a preliminary bating is required. Another disadvantage is that the whole of the enzymes being in the bate their action is more superficial and the grain may be puered too much while the interior of the skin is not penetrated by the enzymes.
The author has made some experiments with an enzyme bate using ammonium butyrate instead of ammonium chloride, and castor oil seed meal in place of the wood fibre.
It was found that the amount of ammonium salts in solution in the most effective bate was approximately one gram per litre. The bating liquid was therefore made up as follows:—
| Ammonium butyrate | 1 | grm. |
| Castor oil seed meal | 2 | " |
| Dry pancreas | 0 | ·01 " |
| Water | 1000 c.c. | |
This liquid at 40° C. bated a skin well in one hour. On increasing the quantity of castor oil seed meal to 4 grm. per litre, the bating effect was not so good. Substituting 0·5 c.c. liquor pancreaticus (Benger) for the solid pancreas, the bating effect was too powerful, the thinner skins were attacked by the bate, and the grain destroyed in patches in a very similar manner to over-puered skins. Further experiments showed that a little more than one-tenth of this amount of pancreatic solution per litre would be sufficient (70/100 c.c. liquor pancreaticus per 1000 litres).
Castor oil seed contains a lipatic enzyme, and when shaken up with olive oil will emulsify the oil. 75 grm castor oil seed meal, and 10 c.c. of olive oil were shaken up with 490 c.c. of water and formed a perfect emulsion, no fat being visible on the surface of the liquid. The mixture was diluted to 7 litres, and a grain treated in it at 40° C. for half an hour; it was fairly well bated in this time, and had a smooth slippery feel, such as skins have in the puer, but was perceptibly “higher” than the skins bated in the new bate containing pancreatin.
This experiment appears to confirm the fact that a mixture of enzymes is necessary, or, at any rate, that a lipatic enzyme alone is insufficient for complete puering.
Dermiforma.142—This is an American preparation, put on the market by the Nowak Manufacturing Co. of Chicago, and is described as a natural bacterial liquor of organic acidity. For 1200 lb. scudded calf skins, 9 gallons of the liquid Dermiforma are used at 100° F., one-third being put in to start with and the remainder added gradually; time required, about 3 hours. For sheep skin a less quantity is required. This bate was found to be a preparation of whey, and contains a certain quantity (18–20 per cent.) of lactic and other organic acids, produced by fermentation; it is therefore to be classed as a drench, and is not a true bate or puer, since tryptic ferments or bacteria are not present.
Puerine.—Another bate, manufactured in America is advertised under the name of Puerine, “the long-looked-for bacterial bate.” The manufacturers give the following directions for use. To start a bating liquor, 10 lb. of the dry powdered puerine is put into a half barrel of warm water (100° F.), and allowed to stand for 3 or 4 days, maintaining the temperature by occasionally warming the infusion with a steam pipe. This amount is sufficient for each 100 galls. water in the bating vat or paddle wheel, and will bate as many skins as can be turned in 100 galls. When the warm infusion of puerine has stood the required length of time, in order to generate the necessary bacterial ferments, the bating vat is run up with warm water say, 100 degrees in warm weather and 110 in cold weather, and the infusion of puerine, including the undissolved portion, is added to the bating vat, and the skins entered into the bating liquor. As the action of the puerine on the skins is exactly the same as a dung bate, the judgment of the operator must determine when the skins are “low enough.” It is not necessary to make a fresh bating liquor for each pack of skins, because the bating liquor improves with age. With each succeeding pack of skins, enough warm water is added to the bating vat to make up for the loss of liquor which is occasioned by the removal of the previous pack, and from 3 to 5 lb. of puerine for each 100 galls. added directly to the bating liquor; it is then in condition immediately to use again.
The bate was found to consist of
| Sugar. | ||
| Gelatin. | ||
| Sodium | Chlorides and sulphates. | |
| Ammonium | ||
| Bisulphite of lime. | ||
| Meal. | ||
As it is allowed to ferment without the addition of bacteria, it will be seen that it acts in quite a different manner to erodin, being more of a modified drench than a true bate, and is not in any way a perfect substitute for the dung bate.
Sucanine.—This is a French bate, put on the market by Messrs. Muller and Co., Rouen; it is described as “concentrated dog puer, chemically clean, i.e., all foreign matters have been eliminated and it contains only that which is really necessary.” The directions state that the quantity to be used is the same as of dog puer prepared ready for use. The paddle is kept at a temperature of 30° C.
This material was found to be a liquid containing ammonium sulphide, phosphates and chlorides of sodium in solution, a considerable quantity of solid matter in suspension consisted of calcium phosphate and calcium sulphide. There was sufficient organic matter to permit of bacteria growing in the diluted liquid.
It had no harmful effect on skins and “brought down” most sheep skins in an effective manner; some resistant skins appeared to require further puering.
Purgatol is a patent bate put on the market by Dr. G. Eberle and Co., of Stuttgart. 1 34 lb. is said to be sufficient to bate 100 lb. wet pelt in 1–2 hours at 38°–40° C. It is a dark brown liquid with an unpleasant smell. When boiled with potash it gives volatile alkaline products.
Distilled with sulphuric acid it gives an acid distillate consisting of organic acids mostly of the fatty series. It contains amines and volatile nitrogenous bases, but no enzymes or bacteria. It must therefore be classed as a deliming agent and not as a true bate or puer.
“Esco.”—This recent artificial bate is of an acid character, and is said to have an action intermediate between the bran drench and pigeon dung; it contains no animal enzymes, but appears to contain some vegetable enzymes of a lipatic character; it also contains 18 per cent. of ammonium chloride in which respect it resembles “Oropon.” It is manufactured by E. Stickelberger and Co., Haltigen, near Basle.
Other artificial bates are treated of in the next chapter under the respective patents. There is also an interesting article on new bates by Eitner in Der Gerber, Nos. 878 and 879, 1911. See also Collegium, 1911, p. 402.
In Chapter VII. we have described a few of the many artificial bates which have been proposed, and for which in many cases patents have been taken out. It would be impossible within the limits of a single chapter to give an adequate account of all these patents, and therefore it is not proposed to deal with those which are merely deliming processes, but solely those in which the use of bacteria and enzymes is directly applied. We mention, however, one or two of the more interesting deliming patents.143
Amend, Otto Paul, New York (U.S. Pat. 763,347, 1904; Eng. Pat. 18,514 and 18,514A, 1904).—Improved Process for Bating and Puering Skins, and Improvements relating to the Bating, De-acidifying and Oxidizing of Hides and Skins.
The skins are treated with a solution of an ammoniacal salt, small quantities of an acid such as hydrochloric acid being added at intervals, so as to reform the salts. Pickled skins are de-acidified and oxidized by treatment in a neutral or slightly alkaline solution of a nitrite of an alkali or alkaline earth such as sodium nitrite.
Nowak, O. H., Chicago (French Pat. 360,854, 1905; Eng. Pat. 26,771, 1905).—Substance for Treating Hides and Process for Making same. This is the “dermiforma” described in Chapter VII. Although bacterial action is used to prepare the bate, the essential action is an acid one, and, as we have said, resembles the drench.
Norris, W. M., New Jersey (U.S. Pat. 840,794, 1907; Eng. Pat. 29,661, 1906).—Improved Process of Treating Hides and Skins. In this process the skins are subjected to treatment with a dilute solution of hyposulphite of soda, which is said to “set” the gelatinous fibres of the skin. The deliming is completed with hydrochloric acid in the presence of salt.
Boehringer Sohn, C. H. (Eng. Pat. 3,140, 1911) Proposes to use in place of the free organic acid, in this case lactic, the sodium salt, and then to add slowly the amount of hydrochloric or sulphuric acid requisite for deliming. The mineral acid combines with the sodium salt, liberating an equivalent amount of the weaker organic acid, which then acts upon the lime in the skins. For deliming 100 kilos. of pelt, 0·25 kilo. of sodium lactate (50 per cent.) and 0·56 kilo. hydrochloric acid (20° Bé.) are used.
The advantage of the method is that a considerable saving is effected in comparison to the quantity of the free organic acid which would be required. In the above example 1 kilo. of lactic acid (43 per cent.) would be required to neutralize a corresponding amount of lime.
In addition to these deliming processes, there are a number of bates in which various nutritive media are allowed to ferment naturally, without the addition of any specific bacteria. Among these may be mentioned:—
Dr. Nördlinger’s Bate, in which a paste of potatoes is allowed to ferment at 30°–37° C. for several days; it is then dried at 40°–50° C., and used in the same way as dog dung.
H. E. Simons’, Phosphorbutyralin, consists of the beetroot residues in sugar manufacture which have undergone an acid fermentation, and contain butyrates and phosphates of ammonia.
Oakes (US. Pat. 798,070, 1905; Eng. Pat. 24,488, 1905).—In this patent an albuminous body, such as casein is allowed to putrefy, at a temperature of 100° F., for several days; an equal weight of flowers of sulphur is then added, and the mixture used as a bate or puer. The sulphur is said to play an essential part in the bate.
A similar process, by the same author (U.S. Pat. 798,293), is described by Dr. Allen Rogers144 in which 5 per cent. of glucose syrup and 10 per cent. of sulphur, computed on the weight of the skins, is fermented with yeast for twenty-four hours, after which it is used as a bate. From a chemical point of view, it is difficult to express the reactions taking place.
When the bath is first prepared, it is neutral in reaction, but, after standing, gradually assumes a slight acid reaction. On introducing the skins, it becomes alkaline, owing to diffusion of the lime. After a short time, it becomes neutral again, and remains thus until the skins are in a perfect condition, when a slight acid reaction is observed. These changes are caused by the fermentation of the glucose, carbon dioxide and alcohol being generated. Owing to the presence of sulphur, a quantity of hydrogen sulphide is produced, which, in its nascent condition, combines with the alcohol to form mercaptans, which in their turn are slowly oxidized to thioacids. These mercaptans and thioacids, being of weak acid character, act in conjunction with any excess of hydrogen sulphide upon the lime, converting it into soluble compounds, which are readily removed by diffusion.
Lederer, L. (French Pat. No. 404,926 of 1909).—Method of Preparing a Substitute for Animal Excrement (dog dung) in Puering Skins for Glacé Leather. In this process a mixture of 10 parts finely powdered bone meal, and 1 part of crushed lupin seeds or beans is formed into a paste with water and allowed to undergo spontaneous fermentation from 3–4 weeks. The mixture is said to form a perfect culture medium for the micro-organisms indispensable in bating. The removal of the lime is facilitated by the addition of 2 per cent. of fat, 0·75 per cent. sodium polysulphide and 1 12 per cent. common salt reckoned on the weight of the bone meal.
In this patent the researches of Wood, Andreasch and Eitner are referred to, and also Benker’s process of bating with Peruvian guano to which soda has been added.
Turney, Sir J. and Wood, J. T.—Coming now to bacterial and enzymatic bates, the author in conjunction with Sir John Turney applied for a patent for a new and improved method of bating skins (No. 25,894, 1896), in which cultures of suitable bacteria were to be used in place of infusions of dog or pigeon dung. It was found however that this patent had been anticipated by the patent 21,720 of 1895, of Dr. Popp and Dr. Becker of Frankfort o/M, and the application was therefore not proceeded with. As has been related in Chapter VII. the late Franz Kathreiner, who knew of the work which was being done both by Dr. H. Becker and the author, made us personally acquainted, and most of our later work was done in conjunction. Like many other new ideas, it was arrived at quite independently by Dr. H. Becker and the author. (See Report of Third Conference of I.A.L.T.C., Copenhagen, 1899, Wiss. Techn. Beilage des Ledermarkt, 1899–1900, p. 8.)
We shall therefore give the original patents in full, and afterwards the translation of Dr. Otto Röhms’ patent, and the recent patent of Dr. Eberle, as these cover the whole ground of the bacterial and enzymatic bates up to the present time.
Dr. Popp and Dr. Becker’s English Patent (No. 21,720, 1895). Improvements in Lyes or Liquors for Preparing Hides for Tanning. An important role in the process of manufacture of many kinds of leather is played by the dung liquor or lye, which in particular for calf skins and skivers constitutes one of the necessary preparatory operations for the subsequent actual tanning process, and which has for its purpose to bring the skins or hides, which have been previously swelled by the liming process, into a condition which allows the easy and thorough removal of the lime and lime soap as well as the remaining hairs by the subsequent mechanical treatment, and to obtain a uniform shrinking of the swelled hides.
The process hitherto employed consists in treating the hides or skins for some time (such as for 4 or 5 hours) in a vat provided with a reel, and containing a lye or brine formed of solutions of dog or pigeon or poultry dung.
Generally speaking, the dog dung acts best in some cases, and pigeon or poultry dung in other cases, but no satisfactory and sound explanation has as yet been given as to the nature of the action.
My correspondents have in the first place ascertained that the action of these lyes or brines is the result of bacteriological processes, especially of the products created by the action of certain bacteria, and is certainly not, as it has hitherto been generally assumed, the result of the action of phosphoric acid or other acids.
The examination of the dog, pigeon and poultry dung in bacteriological respect, and of the industrial applicability of all the different results obtained, shows in the first instance that, even after removal of all other ingredients, which, according to the hitherto ruling views, enter into reaction, the result of the treatment remains the same as it had been before the removal. The continued research led to the second result that a great number of micro-organisms play a role in the lye, partially an advantageous, and partially a detrimental one.
Generally speaking, the non-peptonizing bacteria act advantageously, while the peptonizing ones generally have a detrimental effect. In connexion herewith two methods of utilizing these discoveries have been found.
One can either, according to the methods of bacteriology (plate process), remove all the detrimental or even the less useful bacteria, and effect the treatment of the hide or skins by applying a pure culture of one or at least a mixture of the different most effective bacteria; or one can, without employing such a refined process, but still utilizing the said discoveries, apply the dog dung, and in particular the white descriptions which are most effective, or, for certain kinds of leather, the pigeon or poultry dung, subjecting the said dungs to a certain treatment, which, by the use of a suitable cultivating bed, will develop the useful bacteria, while on the other hand, the increase of detrimental bacteria may be prevented or retarded by the addition of materials hindering the development of the latter.
In every case the employment of the process secures greater rapidity of execution, and more reliable and better results than the process heretofore applied.
The leathers obtained are tough and yet softer, and in any case any discolouring, such as is often unavoidable with dog dung, is avoided.
An advantage not to be underrated with the use of germ cultures, is that liquors or cakes having a definite percentage of really effective substance can be prepared, whereas it was heretofore necessary to proceed by way of trials, the result being subject to great uncertainty.
In addition, the dung liquors are subject to changes overnight under the influence of atmospheric changes (thunder-storms) in a most unaccountable manner, owing, as has now been discovered by my correspondents, to the growth of detrimental bacteria which are mixed with the effective ones, and which gain the upper hand over the latter under the change of atmospheric conditions.
The features of the newly discovered useful bacteria are fully described in the following, and experiments have shown that the particular kinds designed with 3, 7 and 12 in the table of bacteria, set forth in the Specification to German Patent No. 86,335 and hereinafter fully described, are those which are of value for the purpose in question; in particular No. 12 is the most effective, and 3 the least effective of these three descriptions.
All three are bacteria to be found in the dog dung, and bacteria No. 3 consists of very small rods rounded at the ends and are exceedingly mobile, while bacteria No. 7, which are similar to the hay bacillus, are slowly-moving rods, and bacteria No. 12 are medium-sized rods having rapid motion.
The prick-culture gives with bacteria No. 3 a very good growth in the depth; from the inoculating prick extend laterally many arms into the gelatin. At the ends of the arms are formed small knots. On the surface is formed a thin white covering. The gelatin is not liquefied.
On gelatin plates, the deep-seated colonies consist of pale yellow round disks, which gradually penetrate to the surface, and there they form circular disks which show at the centre larger spots. On inclined agar-agar there is formed a white superstratum.
With bacteria No. 7, the gelatin is strongly liquefied in the prick-culture. On the surface of the zone of liquefaction there is formed a white skin.
Along the liquefied prick are formed radiations into the solid gelatin. On gelatin plates there are formed rapidly liquefying colonies forming at top a white skin, agar-agar gives a white unequal thin superstratum with branchings.
Bacteria No. 12 grow with prick-culture equally well along the prick; the gelatin is not liquefied. On the surface there is formed a thin shining covering.
On gelatin plates, the deep lying colonies consist of pale yellow circular disks.
Many colonies have tails similar to the surface colonies of the Proteus. During the growth of this bacterium on gelatin plates a strong putrid smell is developed.
On agar-agar this bacterium forms a strong white shining superstratum.
These bacteria grow easily in any decoction of meat, and the flesh particles from the flesh sides of the hides, which otherwise form a waste product of little or no value, may be advantageously employed for this purpose; bran, and other materials known to offer good culture-beds, may be employed instead.
For this purpose a vessel provided with a cover is charged with equal parts of waste flesh, such as is obtained in the tannery, and water, and this is boiled for about half an hour by the direct introduction of steam, after which concentrated carbonate of soda solution is introduced until the reaction is slightly alkaline, after which the decoction is cooled down. As soon as the temperature has sunk to about 35° C. there is added the pure culture obtained by the gelatin plate process (with dilution), described in the works on bacteriology of Koch, Fraenkel, Hueppe and others, and also by the further treatment corresponding to the characteristic peculiarities of the bacteria, and the whole is allowed to stand covered up for a period of from twelve hours to five days, depending on the time of year and the temperature.
The hides or skins are then dipped into the concentrated decoction for a short time, according to the state of development of the bacteria; or the decoction can be diluted and the hides be then immersed therein for a greater length of time.
The further treatment is then carried out in exactly the same way as though the hides or skins had been treated in dog-dung lye, with the exception that the duration is shorter.
In case the pure culture of the bacteria is to be dispensed with, and it is only desired to obtain an advantage over the existing methods in so far as to obtain, on the one hand an artificial increase of the ooze medium viz. the number of useful bacteria and their products existing in the dog dung, by providing a suitable culture bed, and on the other hand to be in a position to limit oneself to the best initial material, the process can be carried out by starting with a practically proved dog dung, therefore in particular the white kinds, and the bacteria are caused to propagate in a decoction such as above described. Afterwards it is advantageous to effect the killing of the detrimental bacteria by a small addition of acid, such as 0·5 per cent. of dilute sulphuric acid or equivalent materials, as in this case the useful bacteria are also the most enduring.
The more useful bacteria show, in particular in the presence of alkalis, the same behaviour.
For example, the more useful bacteria arrive at development equally with the detrimental ones, if to the described culture bed be added 0·5 per cent. of carbonate of soda, while the useful bacteria preponderate to a considerable extent, and the detrimental ones are practically unimportant, if the additions of carbonate of soda be raised to about one per cent.
The pure cultures of the bacteria may be preserved in a suitable liquid or solid material, and brought on the market in a suitable packing.
In order to obtain a product which shall both be durable, and capable of being delivered in definite doses, sterilized bran, flour, etc., can be inoculated with definite quantities of the pure cultures, and the percentage of water be reduced, by drying, pressing, etc., to under a certain percentage (10 to 12 per cent.) which will not allow of, or will retard the evolution of germs.
The bacteria may also, after having been properly developed, especially after having created a sufficient quantity of their products, be killed, for instance by heating, and the resulting stuff be brought on the market in liquid or solid form, as an effectual lye or brine, it having been found that the liquids containing the said bacteria do not lose their effect if they are sterilized and the bacteria thereby killed, provided that a sufficient number of bacteria has been produced.
Claim.—1. The process for the production of a lye for treating hides or skins preparatory to tanning, consisting in the propagation of the bacteria of dung, in particular dog and bird dung, by means of suitable culture beds substantially as described.
2. As a new article of manufacture a lye containing a pure culture of one or more of the described bacteria.
3. In the process of producing a lye referred to in the first claim, the elimination of the detrimental agents substantially as described.
4. In the process referred to in the first claim, the addition of acids or alkalis for the purpose set forth and substantially as described.
5. The process for the production of a lye for treating hides, consisting in the separation from the dung of the bacteria herein designated as 3, 7 and 12, producing pure cultures thereof, and introducing the same into a decoction preferably freed by boiling from other bacteria and containing a suitable culture bed, substantially as described.
6. A process for the production of cakes, for the treatment of hides, consisting in the inoculation of sterilized bran, flour, etc., with cultures having a definite percentage of bacteria and the removal of the proportion of water that is necessary for the development of the bacteria, substantially as described.
7. As a new article of manufacture sterilized cakes, containing a certain quantity of the described bacteria, substantially as described.
8. As a new article of manufacture the solid or liquid material containing the products of the decomposition, as generated by the pure cultures of the said bacteria, whether such material does or does not also contain the said bacteria, either in the active condition or killed by sterilization, substantially as described.
Wood’s English Patent (12,549, 1898).—Improvements in and relating to the means used for the Bating of Hides and Skins. This invention has reference to improvements in and relating to the means, liquid or bate, used for the bating or puering of hides and skins previous to the process of tanning the same.
At the present time skins are usually bated by means of a liquid or bate prepared from dog or pigeon dung, and although the bating action or actions have not hitherto been thoroughly understood, it is a well known fact that fermentations going on in the dung play an essential part. It was also pointed out in 1885 by H. R. Procter, in his “Text Book on Tanning,” that the action of the bate was in a great measure due to bacteria. It is also a known fact to practical tanners that a dung bate, properly applied, produces the best quality of leather.
Owing, however, to the varying composition of the dung used, the bate made therefrom possesses varying properties, its bating action is uncertain, its properties are readily affected by atmospheric influences, and the skins treated are liable to putrefaction.
As a substitute for dung bate, it has been proposed, as set forth in the Specification of Patent No. 21,720, A.D. 1895, to use a bate made from a pure culture of bacteria. Such bate, however, does not possess all the essential properties of a dung bate; and although bacterial action is necessary, such action alone is not sufficient, as was pointed out by the present inventor in 1894 in the “Journal of the Society of Chemical Industry.”
The present inventor has now discovered that the bating action of a dung bate is due to the combined action of enzymes or unorganized ferments, and to certain amine compounds in the dung. These compounds consist of amines (i.e. ammonia in which one or more atoms of hydrogen has or have been replaced by alcohol radicles), in combination with lactic, acetic and other organic acids contained in the dung, the acids named being the principal ones.
The present inventor has also discovered that the bating action of the enzymes alone, or the action of the chemical compounds alone, are inefficient, and that the enzymes exert their bating action in the presence of the chemical compounds, while these latter in addition have an independent action.
The object of this invention is the production, according to scientific methods, of a bate having similar properties to those of a dung bate, such properties in the improved bate being defined and definite, but capable of being modified according to the requirements of the skins to be bated and the leather to be produced.
According to this invention, the liquid or bate is made by producing by fermentation an enzyme or enzymes of the same character as the enzymes contained in dog dung or produced from bacteria obtained from dung, and to the liquid thus obtained there is added an organic acid and an alkali.
The inventor has discovered that for an efficient bate a pure culture of bacteria is insufficient, and that no single organism hitherto isolated will give results equal to dung. He has also discovered that the production of these enzymes depends more upon the composition of the nutrient medium in which the bacteria are cultivated than upon the kind of bacteria, although these latter must be capable of producing the required enzymes, the production of which appears to be due to a symbiotic growth of bacteria, at present little understood.
It is desirable that the nutrient medium should be without sugars or starchy matters, and, although the composition of the nutrient medium may be varied, good results can be obtained according to ordinary bacteriological methods by using a medium composed of—
| Gelatin | 50 | parts |
| Potassium or sodium phosphate | 1 | do. |
| Water | 2500 | do. |
This medium is maintained at a temperature of about 37° C. (the most favourable temperature for the fermentation) until the required bacterial action is completed, the time necessary being about seven days, and to each litre of the liquid thus obtained there are added from 2 to 6 grs. of lactic acid. Acetic or other similar organic acid may be used instead of the lactic acid, but sulphuric or mineral acids must not be used. The acid liquid is afterwards neutralized by ammonia, which is added when the bate is to be used, and there are then formed certain chemical compounds similar to those found in a dung bate.
The addition of the acid to the nutrient medium will immediately stop the growth of bacteria, but the acid is not added for that particular reason, but for the particular purpose set forth.
Instead of the enzymes obtained from the bacteria of dung, other enzymes of a similar character may be used, such as the enzymes obtained from the bacteria found on skins during the sweating process.
For bating skins in a practical manner with the improved bate herein set forth, the method of using the same is preferably varied according to the skins to be treated and the kind of leather it is required to produce. Thus for kid skins, skivers and other skins for which dog dung has hitherto been used, the skins, after the liming process, are first well washed in water in the usual way to remove the bulk of the lime. The skins are then placed in a wheel or paddle with the bate prepared as before described, with the acid exactly neutralized by the ammonia or so nearly neutralized that the alkaline action of the skins will complete the neutralization, all skins being alkaline when introduced into the bate.
The bate is diluted according to the amount of liming the skins have undergone, and when in the paddle or wheel the skins and bate are maintained at a temperature of about 37° C., and the skins paddled until they have “fallen” considerably. The skins are then removed from the wheel or paddle and scudded preferably on the “grain” side, after which they are returned to the wheel or paddle and the paddling continued until the bating action is completed, this being judged in the usual way by a practical tanner according to the feel of the skins. The strength of the bate after the scudding process may be varied if found desirable.
After the bating process is completed the skins are treated in the usual manner.
When the improved bate is to be used for light hides and similar skins which have hitherto been treated with pigeon dung, the skins, after the liming process, are washed in the usual manner, heated to a temperature of about 37° C., and placed in a wheel or paddle containing by preference the improved bate in an acid condition, that is before the ammonia has been added, the bate being previously diluted according to the amounts of liming the skins have undergone. The skins are then paddled until the tanner, by the feel, judges that the whole of the lime has been removed. The acid bate is then neutralized by ammonia, with or without removing the skins, and the paddling is continued until the bating process is completed, this being judged of by the feel of the skins.
The bated skins are then scudded, washed, and treated in the usual manner.
The strength of the bate, into which the skins are placed for both methods, is varied according to the amount of liming the skins have undergone; that is to say, for “high” limed skins, a stronger bate is required than for skins which have been subjected to a moderate liming.
Claim.—1. A bate having the essential properties of a natural dung bate, but prepared according to scientific methods from the means and in the manner substantially as set forth in the accompanying specification.
2. The bate described and set forth in the accompanying specification.
Wood’s American Patent (638,828, Dec. 12, 1899) Bate.—This invention has reference to improvements in and relating to the means, liquid, or bate, used for the bating or puering of skins previous to the process of tanning the same.
At the present time, skins are usually bated by means of a liquid or bate prepared from dog or pigeon dung; and, although the bating action or actions have not hitherto been thoroughly understood, it is a well-known fact that fermentations which the dung undergoes play an essential part. It was also pointed out, by H. E. Procter in 1885, in his “Text Book of Tanning,” that the action of the bate was in a great measure due to bacteria. It is also a known fact to practical tanners that a dung bate properly applied produces the finest and most supple leather. Owing, however, to the varying compositions of the dung used, the bate made therefrom possesses varying properties: its bating action is uncertain, its properties are readily affected by atmospheric influences, and the skins treated are liable to putrefaction.
As a substitute for a dung bate, it has been proposed, as set forth in the specification of the British Patent No. 21,720, A.D. 1895, and the specification of the German Patent No. 86,335, Class 28, to use a bate made from a pure culture of bacteria. Such bate, however, does not possess all the essential properties of a dung bate, and, although bacterial action is necessary, such action alone is not sufficient, as was pointed out by me in 1894 in the “Journal of the Society of Chemical Industry.”
I have now discovered that the bating action of a dung bate is due to the combined action of enzymes, or unorganized ferments, and certain chemical compounds, which compounds consist principally of amines—i.e. ammonia in which one or more atoms of hydrogen has or have been replaced by alcohol radicles—in combination with organic acids usually found in dung, and of the compounds so formed the lactates and acetates are the most important. I have also discovered that the bating action of the enzymes alone, or the action of the above-referred-to chemical compounds alone, is inefficient, and that the enzymes exert their bating action in the presence of the chemical compounds, while these latter, in addition, have an independent action on the limes in the skins, and on the skin fibre.
The object of this invention is the production, according to scientific methods, of a bate having similar properties to those of a dung bate, such properties in the improved bate being modified according to the requirements of the skins to be bated and the leather to be produced.
According to this invention, the liquid or bate is made by producing by fermentation, an enzyme or enzymes of the same character as the enzymes contained in dog dung, or produced from bacteria contained in the dung, and to the liquid thus obtained there is added an organic acid and an alkali.
I have discovered that for an efficient bate a pure culture or single species of bacteria is insufficient, and that no single organism hitherto isolated will give results equal to dung. I have also discovered that the production of enzymes of the character above referred to, depends more upon the composition of the nutrient medium in which the bacteria are cultivated, than upon the kind of bacteria, although these latter must be capable of producing the required enzymes, the production of which appears to be due to a symbiotic growth of bacteria at present little understood. The bacteria used for the purpose of this invention may be the bacteria found in dog dung, or the bacteria obtained from the roots of the hairs of skins during the sweating process, these latter being used, by preference, on account of the ease with which they are obtained, compared with the difficulty of isolating from the dog dung the particular mixture of species most desirable, there being so many undesirable species present. Of the bacteria found on the roots of the hair during the sweating process, and hereinafter referred to as “sweating” bacteria, I have found that two species predominate, and, so far as I am aware, these have not been previously isolated. The principal organism, which I call “bacillus d,” in continuation of a series of previous investigations described in the “Journal of the Society of Chemical Industry,” A.D. 1898, forms large whitish colonies, with irregular contour spreading on the surface of the gelatin, the bacilli being very small, mostly occurring in pairs, but sometimes joined together in thread-like forms. The second organism, called by me “bacillus e,” forms small brownish-yellow boat-shaped colonies on gelatin plates. The bacteria consist of plump cells two or three times the size of bacillus d, united in pairs and chains, but the cells vary considerably in size, and are surrounded by a capsule. I have discovered that the above bacteria, when used separately as pure cultures, exert little or no action on the skin, but when used together the action is very remarkable.
The gelatin and gelatin plates referred to in the above description are the gelatin and gelatin plates used for the cultivation of bacteria, according to the ordinary bacteriological methods as described in Günther’s “Bakteriologie,” published by George Thieme, Leipzig, in 1898, and in other text-books.
In the accompanying drawings, made from photographs which are of record in this case, the bacillus d is shown in Fig. 21, and the bacillus e in Fig. 22, both photographs showing the bacilli magnified 1000 diameters.
A culture may be prepared from the sweating bacteria by taking the hair from the skin when it “slips”—that is, when it can be removed by simply pulling it. The root portion of the hairs are then cut off, and about 10 grm. carefully digested in 100 c.c. of water at a temperature of 32° to 35° centigrade for several hours. The liquid portion is then poured off, and used for inoculating 10,000 c.c. of a nutrient medium of the character hereinafter described. The inoculated nutrient medium is then maintained for three days at a temperature of 37° centigrade, and may then be used for the inoculation of a larger quantity of nutrient medium.
Theoretically, a single cell of a bacterium is sufficient to inoculate any quantity of any suitable fluid; but in practice it is found necessary or expedient to employ 5 or 10 per cent. of inoculating matter to the volume of the liquid to be inoculated. The inoculation of the 10,000 c.c. of nutrient medium may be done in a Carlsberg vessel, as described by Hansen in his “Untersuchungen aus der Praxis der Gärungsindustrie.”
It is desirable that the nutrient medium should be without sugars or carbohydrates; and, although its composition may be varied, good results can be obtained by ordinary bacteriological methods, by using a medium composed of gelatin, 50 parts or 20 grm.; potassium or sodium phosphate, 1 part or 0·4 grm.; sodium chloride, 1 part or 0·4 grm.; water, 2500 parts or 1000 c.c. This medium, after inoculation, is maintained at a temperature of 37° centigrade, until the required bacterial action is completed, i.e., until the whole of the nutriment for the bacteria is exhausted, at which time the maximum quantity of the required enzymes has been produced. The time required for this action is not less than three days, nor more than seven days. The exhaustion of the nutriment may be ascertained scientifically by a microscopic examination of the liquid, the bacilli at this period commencing to form spores. To each litre of this liquid there is then added from 2 to 6 grm. of lactic acid, and the acid liquid is afterward neutralized by ammonia, which is added when the liquid is to be used as a bate, the addition of the ammonia forming certain chemical compounds similar to those found in the dung. Instead of lactic acid, acetic or other similar organic acid may be used, but sulphuric or mineral acids must not be used. The addition of the acid to the liquid will immediately stop the growth of the bacteria; but the acid is not added for that particular purpose, but for the formation of the chemical compounds above referred to.
The above-described method is most suitable, when the bate has to be kept for a time before being used, or when it is desired to concentrate it for transit or export; but if the bate is to be used immediately, it may be prepared as follows: 100 parts of gelatin are dissolved in 1000 parts of water, and there is then added 50 parts of commercial lactic acid. The mixture is then heated, in a closed vessel, to 100° centigrade, by which operation the gelatin is partially peptonized, and the subsequent bacterial action is hastened. The acid liquid is afterward neutralized with ammonia, or other alkali, and the whole diluted with water to 20,000 parts. The dilution may conveniently take place in the wheel or paddle in which the bating is to be done, and to the liquid thus obtained there is added from 5 to 10 per cent. of the active culture from the Carlsberg vessel, as above described. The liquid is then allowed to stand for 15 to 20 hours at a temperature of 37° centigrade, and is then ready for use.
For bating skins in a practical manner with the improved bate herein set forth, the method of using the same is varied, according to the skins to be treated and the kind of leather it is required to produce, and the strength of the bate is varied in exactly the same manner as the strength of a dung bate, and such variation is made according to the judgment of the tanner, the improved bate acting in the same manner as a dung bate.
For kid-skins, calf-skins, skivers, and other skins for which dog dung has hitherto been used, the skins, after the liming process, are first well washed in water in the usual way, to remove the bulk of the lime. The skins are then placed in a wheel or paddle, with the bate prepared, as above described, with the acid exactly neutralized, or so nearly neutralized, that the alkaline action of the skins will complete the neutralization, all skins being alkaline when introduced into the bate. The bate is maintained at a temperature of 37° centigrade, and the skins are kept in motion until they have “fallen” considerably, i.e., until the swollen condition of the fibres due to the liming has disappeared. The skins are then removed from the wheel or paddle and scudded by hand or machine, preferably on the grain side, and the skins not sufficiently reduced are returned to the paddle, and the bating continued until the action is completed, this being judged in the usual way by the tanner by the feel of the skins. The strength of the bate, when skins are returned thereto, may be varied from the original strength, if desired, and after the bating is completed the skins are treated in the usual manner.
The strength of the bate into which the skins are placed, is varied according to the amount of liming the skins have undergone—that is to say, for a high-limed skin a strong bate is required. For moderately-limed skins the bate may be of such a strength that each litre as prepared for use contains five grm. of the original gelatin contained in the nutrient medium, which is equal to one part of liquid in which the culture has taken place to three parts of water. When the bate is prepared for immediate use, its strength is varied in the paddle as required. For very hard skins, such as goat-skins, the culture may be used as a bate undiluted or it may be diluted with an equal quantity of water.
For light-dressing hides and similar skins, which have hitherto been treated with pigeon dung, the skins are washed in the usual manner after the liming process, heated to 37° C., and placed in a wheel or paddle containing the improved bate, preferably in an acid condition—that is, before the ammonia or alkali has been added—the strength of the bate being equivalent to five grm. of original gelatin contained in the nutrient medium to each litre of the bate used. When the lime has been removed from the skins by the bate, which will be in about one hour, a quantity of the original culture previously neutralized with ammonia or other alkali and equal to about one-half of the quantity of the original culture first added is placed in the wheel and the bating continued until the process is completed, this being judged by the feel of the skins. If in the above method the chemical reaction of the bate be examined before the second portion of culture be added, it will be found neutral, and the action of the enzymes takes place in a neutral or alkaline solution, as in the first-described method.
The character of the principal enzyme in dung and in the improved bate is a proteolytic one; i.e. it resembles the trypsin contained in pancreatic juice.
Instead of adding an organic acid and neutralizing it by an alkali, as described with reference to both processes, there may be added an organic salt of the same character as that formed by the combination of the acid and the alkali.
Claim.—1. A bating liquor or bate containing enzymes of the same character as the enzymes contained in dog dung, an organic acid, and an alkali, or their chemical equivalent, i.e. an organic salt substantially as herein set forth.
2. A bating liquor or bate containing enzymes of the same character as the enzymes contained in dog dung and produced by fermentation an organic acid and an alkali, or their chemical equivalent substantially as herein set forth.
3. A bating liquor or bate containing one or more enzymes of the character of the enzymes contained in dog dung, an organic acid and an alkali, or their chemical equivalent substantially as herein set forth.
4. A bating liquor or bate containing one or more enzymes of the character of the enzymes contained in dog dung, or produced from bacteria obtained from dog dung, an organic acid, and an alkali or their chemical equivalent substantially as herein set forth.
5. A bating liquor or bate containing enzymes of the same character as the enzymes contained in dog dung, two to six grm. of an organic acid to each litre of the liquid and neutralized by an alkali substantially as herein described.
6. A bating liquor or bate containing enzymes of the same character as the enzymes contained in dog dung, from two to six grm. of lactic acid to each litre of liquor and neutralized by ammonia substantially as herein described.
7. A bating liquor or bate containing enzymes of the same character as the enzymes contained in dog dung and obtained from bacteria found on the roots of hairs during the sweating process, from two to six grm. of lactic acid, and neutralized by ammonia substantially as herein set forth.
8. A bating liquor or bate containing enzymes produced from bacteria obtained from dog dung and cultivated in a medium composed of fifty parts of gelatin, one part of potassium or sodium phosphate, one part of sodium chloride and two thousand five hundred parts of water, two to six grm. of lactic acid to each litre of liquor and neutralized by the addition of ammonia substantially as herein described.
Oropon (German Patent 200,519, July 21, 1908).—Process for Bating Hides. Dr. Otto Röhm, Esslingen a/Neckar. In the present process, a watery extract of the pancreas in combination with ammonia or alkaline salts or mixtures of these salts is used as a bate liquor. The object of the process is to replace the dung bate. From the researches of Wood145 also Jettmar146 it appears that the peculiar behaviour of the dog dung bate is to be attributed to the action of enzymes in combination with organic amine compounds and ammonia salts. Starting from the assumption that other enzymes, known to have an action on hide substance, might have an analogous action to the dog dung bate, the author discovered that the trypsin of the pancreas had exactly the same bating action as the enzymes of dog dung. The action is aided by the fat-splitting enzyme of the pancreas, steapsin, which saponifies the fat in the hides.
A good bating action is obtained by a watery extract of pancreas, by the addition of an ammonium salt, which changes the caustic lime (from the limed pelt) into a soluble lime salt, and gradually reduces the alkalinity of the bate liquor caused by the lime in the skins, since the ammonia liberated by the lime escapes to a great extent from the liquor. The favourable action of the ammonium salt on the skins causes them to fall and become thinner, nor do they become rough when placed in clean water after bating, as a strongly alkaline skin will if placed in water containing calcium carbonate.
For example, the process may be conducted by extracting a pancreas weighing about 250 grm. with 1 litre of water, and adding 10 c.c. of such extract to 990 c.c. of a solution containing 0·15 per cent. ammonium hydrosulphide and 0·3 per cent. sodium chloride. Such a solution forms a very active bate. In place of the ammonium hydrosulphide, any other ammonium salt yielding a soluble lime salt may be used, e.g., ammonium chloride.
The pancreas extract used in the above process must be used in a fresh condition or preserved from putrefaction by the addition of some suitable preservative; if putrefied, and consequently acid, it will not work. The preservation of the pancreas may also be brought about by drying, and the bate liquor made from the dry material by putting it in water. No putrefaction occurs during the bating process. The action of the enzymes is favoured by the alkaline reaction of the hide.
Claim.—Process for bating hides, characterized by the use of a watery extract of the pancreas, with the addition of an ammonium or alkali salt which forms soluble lime salts, or mixtures of such salts to the liquid.
In an addition to the above patent, Dr. Röhm proposes to soften the water used by adding the proper amount of clear lime water to it. Starch paste can also be used for the same purpose, or a mixture of lime water and starch paste.
In a further addition, a weak acid mixture, consisting of 0·05 to 0·1 per cent. lactic acid and 0·05 to 0·1 per cent. ammonium chloride, is proposed in place of the ammonium salt alone, since the evolution of free ammonia had been found to be detrimental to the skins.
Eberle’s English Patent (21,202, 1909).—Improved Process for Bating Hides. Efforts have been made for years to replace dogs’ excrement used in many cases in tanning as a bate, by other simpler products and methods which in particular shall have the advantage of greater uniformity and thereby act with greater certainty.
Now it has been ascertained that substances which are only solvents of lime, such as acids of inorganic or organic nature and their salts, form an incomplete substitute. It has been shown by Eitner, Wood and others that there are enzymes and bacteria to which dogs’ dung owes its specific or bating properties. The action of dogs’ dung is explained as one which liquefies or dissolves the skin substance, such as is otherwise only done by enzymes and bacteria.
Experiments in the laboratory and in practice have shown that it is not the dissolving action on the skin substance in the narrow sense which is desired in bating the derm with dogs’ dung, but in order to prepare the hide for the manufacture of a soft and elastic leather, the albuminous intercellular substances and more particularly the layers of fat lying at the roots of the hair, in the corium and in many cases also on the epidermis, must be dissolved and thereby the fibrils of the skin loosened. The highly solvent action of the dung bate on the skin substance means in very many cases a direct disadvantage by reason of the loss of leather substance.
Extensive experiments with the separate constituents of dogs’ dung have shown that with none of them employed alone can a satisfactory action be obtained. For the experiments, the organic and inorganic salts contained in dog’s dung were employed. Pure cultures of intestinal bacteria were also examined and also the intestinal juices, secretions of the gall bladder, of the pancreatic gland and secretions of the intestinal mucous membrane. The relatively best bating action was obtained with the secretion of the gall bladder; only a moderate bating action could be obtained with the juices of the pancreatic gland and the secretions of the intestinal mucous membrane. It was therefore very surprising to find and could not be foreseen that mixtures of these three secretions of the digestive apparatus alone exerted an excellent bating action, and that such mixtures not only formed a perfect substitute for the bate of dogs’ dung, but that they acted far better than the latter, without it being necessary, as was recommended elsewhere,147 to add lime-dissolving salts to the bate, or to take measures in any other manner for the removal of the lime from the water.148
The explanation of the effective action of such mixtures is evident from the following statements:—
Pancreatic juice contains three ferments, of which for our purpose trypsin, which decomposes albumen, and steapsin which decomposes fat, are the most important. Both ferments are contained as zymogens, that is to say in an inactive form, in the pancreatic juice. The latter therefore only gives an incomplete bating action. Gall contains ferments which have properties which are mainly fat decomposing, but are also albumen dissolving. Gall also contains the salts of gallic acids which have the power in a high degree of dissolving and emulsioning greases or fats, and among others have a great solvent action on the chloresterin contained in the skin, more particularly in the roots of the hair. As regards the fat-decomposing and dissolving action, gall is far superior to pancreatic juice.
The ferments of gall are, however, also capable of energising the action of the steapsin in the pancreatic juice as a fat-decomposing ferment, that is to say of imparting to the pancreatic juice, which in a pure condition has a very slight action on fat, a high capacity for decomposing fat.149
Finally gall in bating the derms may be of great importance inasmuch as its acids form easily soluble lime salts. Extensive experiments have shown that gall, as such, is adapted to dissolve a comparatively large quantity of lime. In hard water an addition of gall prevents the precipitation of calcium carbonate, and the roughening of the derm, which, for instance, has been noticed when pure bacteria cultures are employed, is prevented by the addition of gall.
The intestinal juice contains in addition to carbonates and alkalis, enzymes which again exert a slighter bating action on the hide than those of gall. The intestinal juice alone has a similar action to pancreatic juice, but if intestinal juice be added to pancreatic juice, the most powerful enzyme of pancreatic juice, namely the albumen-decomposing trypsin is energised, and only in this way does the pancreatic juice fully acquire the property of dissolving albuminous substances.150
The proportions in which such mixtures best develop their actions, are those which obtain in animal organisms; the temperature also especially plays a great part. The maximum temperature lies about 40° C.
From the above statements it is evident that it is possible to produce various actions with the juices of the pancreas, gall and the intestines, quite dependent on which are considered the most important in the bating. Similarly, as the proteolytic action of the mixture of the three juices may be increased or diminished by using larger or smaller quantities of intestinal juice and pancreas, the fat-decomposing and fat-dissolving action of the mixture may also be increased or diminished by using larger or smaller quantities of gall, and of gall and pancreas respectively. Finally the quantity of the gall to be added is decided according to the amount of lime contained in the water employed in the bating. Experience has shown that a relatively small addition of gall will act very advantageously.
It must be especially remarked that artificial bates made in this way are extremely cheap as compared with the natural dogs’ dung, as gall and intestinal juice may be procured at very low prices and a considerably smaller quantity of the energised, more expensive, pancreatic juice is necessary than, for instance, would be necessary if this juice were employed in a pure condition.
The advantage of this invention also consists in its being possible to make better bates than that which is made, for instance, from dogs’ dung because, as hereinbefore stated, it is a disadvantage of this latter bate that it in many cases dissolves too large a quantity of the albuminous substance. Otherwise, the composition of the improved bate quite coincides qualitatively with the composition of dogs’ dung, as the latter actually consists in addition to the unabsorbed portions of food, of secretions of gall, the pancreatic gland and the mucous membrane of the intestines. The trypsin and steapsin respectively as well as being derived from the pancreatic juice, may also be derived from the vegetable kingdom, for instance, there are considerable quantities of trypsin in the sap of the fig. Steapsin, in not inconsiderable quantity, is found in the saps of flax, hemp, maize, etc.
It must also be remarked that the colouring matters in gall are of quite subordinate importance, and may be disregarded. The view formerly adopted that the colouring matters in gall stain the leather is incorrect.
Example:—200 c.cm. of gall juice in 1 litre of water are mixed with the watery extract of 200 grm. of chopped small intestine in 1 litre of water on the one hand, and 100 grm. of the pancreatic gland in 1 litre of water, on the other hand. This watery solution is added to the bating bath, according to the bating action desired. It can also, in order to allow of being more easily handled, be obtained in solid form by absorption in sawdust or kieselguhr and drying at 30° to 40° C.
Claim.—A process for bating hides, characterised by the use of mixtures of gall, the secretion of the pancreatic gland and intestinal juice, and by the use of mixtures of the active enzymes in these, or active enzymes derived from the vegetable kingdom, substantially as described.
Prof. Dr. H. Becker’s English Patent (24,982, 1910).—Improvements in Puering and Bating Hides and Skins.—It has been shown by Wood, of Nottingham, and by Dr. Popp and Dr. Becker, of Frankfort-on-the-Main, that the action of a good puer or bate on hides, is connected with the development of certain species of bacteria.
The preparation of pure cultures of these bacteria (which are derived from dog dung) has enabled what may be properly termed a safe puer to be obtained, inasmuch as by its use overpuering the hides is entirely prevented. Furthermore, the leathers are fuller and do not lose any excess of hide substance, so that a dozen sheepskins that have been suitably puered by the bacteriological method, will weigh about 9 ounces more than a dozen skins of the same kind and origin, that have been treated in exactly the same way in other respects, but puered with dog dung.
The method of action of the bacterial puer has been elucidated as follows. The bacteria present in the puer are microscopically small and pass easily into the skin through the pores of the latter, while on the surface and in the substance of the skin they produce their metabolic products and enzymes. These substances are partly proteolytic, partly fat decomposing, and also diastatic in character, so that the highly proteid intermediate substance of the skin is degraded. By means of the ultimately formed ammonia, amine bases and ammonium bases assisted by the action of organic acids produced by the bacteria, the lime soaps are decomposed. Lime and fat are partially dissolved, and partially further decomposed or converted into a form which enables them to be worked out by mechanical means. At the same time the roots of the hairs are also loosened so that the hairs also can be readily removed by mechanical treatment.
Another essential factor is the liberation of gases formed within the skin by the action of the micro-organisms and exerting a very gentle pressure on their environment. The structure of the fibres is loosened, the fibrils are straightened individually without sustaining any important loss of corporeal matter, and in this way a firm, tough and yet soft handling leather is obtained. Attempts have been made to improve this puer by adding, for instance, organic acids and salts to the bacterial cultures.
Other methods of puering, based on the direct weakening of the skin fibres from the outside, may also effect the pulling down of the hides, and moreover, in consequence of that attack on the skin fibres without straightening the fibrils, an immoderate amount of substance is lost, and therefore a hard, stiff, thin and light leather is obtained.
To this class belong also the purely digestive puers such as pancreatic juice, intestinal juice and the like, or purified enzymes. These enzymes are colloidal and therefore incapable of diffusion, and consequently they digest the hide mainly from the surface, that is to say, the grain side and inner side of the hide. The fibres, in-so-far as they are accessible at all to the action of the colloidal enzymes, are removed from outside without the fibrils being loosened and straightened. Hence they are weakened by the digestive puer to such an extent that the hides seem to be gradually pulled down; but the skin will by that time have lost too much substance, so that, inter alia, even the grain appears to have been corroded. In this way the leather is rendered thinner and lighter than that from the dung or bacterial puer—a defect manifest to every expert.
This danger is particularly great when, by the artificial admixture of corporeal juices, complications are set up, that intensify the destructive action of the digestive enzymes on the material, and when their supplementary action is not suitably controlled.
These animal or digestive enzymes and fat decomposing substances originate, in-so-far as they are not of vegetable origin, for the most part in the upper portions of the digestive tract. The use of these substances effects an imitation of the purely enzymatic and fat decomposing processes that go on in the upper part of the intestines, but omits the processes—not less important for digestion and the formation of excrement—occurring in the large intestine and the adjoining portions of the digestive tract. Here the bacteria and the vegetable enzymes secreted thereby, play the chief part. They decompose not only the food constituents—partly by putrefaction—but also the albuminoid digestive secretions and the gall, so that only small quantities thereof are left. Hence when applied to the puering process, the use of bacteria, together with their specific enzymes and metabolic products, is equivalent to a restriction of the immoderate digestive action of the pancreatic and digestive juices, i.e. of the animal enzymes.
Digestion therefore, with the resulting formation of excrement, is carried on with the accompaniment of a mutual control, excitation and restriction on the part of the digestive secretions and the bacterial phenomena.
These latter form the real basis of the puering of hides in dung preparations.
This is also evident from the purely practical experience that dog dung puer, e.g. for glove leather, is essentially improved by leaving the dung to ferment for several weeks or rather months. During this process an increase occurs in the micro-organisms present, and also in their metabolic products and enzymes, but not in the digestive juices originating in the organs of the animal body—the term “digestive juices” being restricted to indicate only such juices as are produced in the course of the digestive tract by the organs and cells of the animal body, in contradistinction to the juices produced by bacteria.
According to the present invention, the bacterial cultures, together with their metabolic products or products of digestion and enzymes, are used in combination with one or more of the substances that are excreted in the digestive tract by the animal body itself in order that not only each of these substances may exert its special action on the skin, but also that the aforesaid mutual influencing, and more particularly regulating of the action may take place.
On this principle, in preparing an artificial puer for hides, and in accordance with the nature and preliminary treatment of the raw material, and the requirements put forward in respect of the finished product, the bacterial puer, nutrient medium or culture is mixed with the corresponding digestive juices, or with purified animal or vegetable enzymes, as well as with fat decomposing media, if necessary, such as ground castor-oil seeds, hemp seed or the like, or with extracts of same, or with gall. Where it seems necessary, substances capable of dissolving lime may also be added, for example, sugar, molasses, salts of ammonia, organic acids, or the corresponding salts or the like.
Finally, also, the several operations may be carried out independently of one another, it being in such case again advisable to adhere to the processes going on in the intestines.
If, for instance, specially hard goat skins are to be pulled down strongly by puering, it is advisable to add to the nutrient medium of the bacteria a little papayotin or papain, either—according as rendered necessary by the local conditions—in the preparation of the bacterial puering liquor, or during the propagation of the bacteria. If the fells are less hard so that less pulling down is required, the hides are first treated with a weak solution of the digestive ferments, and then with the bacterial liquor. In this way excessive digestive action is counteracted in good time.
Sheep and goat skins that are particularly rich in fat are immersed in a bacterial puering liquor, containing ground castor-oil seed or castor-oil cake in suspension. The fat-decomposing ferment contained in this meal then comes into immediate action, and the liberated fatty acids form readily soluble soaps with the ammonia, originating from the degradation of the proteid substances, and with the allied bases.
Extensive practical experiments have also shown that, in the case of very fat skins, the action of the bacterial puer is increased by mixing a suitable salt of sulpho-ricinoleic acid with the puering liquor, or by washing the skins with solutions of similar soaps either before they are immersed in the puering liquor, or subsequently.
Local conditions, and the requirements laid on the finished leather, must decide which of these methods is specially advisable.
In any event, however, the combinations of the bacterial puer afford a means of obtaining an increased puering effect in many cases where puering with the separate auxiliaries mentioned does not produce a sufficient result.
Claim.—In the method of puering hides wherein bacterial cultures, their metabolic products and enzymes are employed, the combined use with these agents of digestive juices, mixtures of digestive enzymes or pure digestive enzymes (and, if desired, of fat-decomposing substances), either simultaneously with said agents or subsequently thereto.
In the manufacture of many kinds of light leathers, skins, after bating, are drenched. The process usually consists in placing the skins in a mixture of bran and water, half to one per cent. of bran being used (5 to 10 grm. per litre, or 12 to 1 lb. per 10 gall.) at a temperature of 29° to 35° C. This ferments vigorously for 18 to 24 hours, with evolution of a considerable quantity of gas, and the formation of weak organic acids.
The acids neutralize any lime which has not been neutralized in the bate, and they do this in a very gradual and mild way, being produced by bacterial fermentation exactly as required, in a similar manner to fermentations taking place in presence of calcium carbonate. The gases evolved bring the skins to the surface of the drench, and distend the fibres; the workman attending to the drenches puts the skins down under the liquor with a pole, but a better way is to have the drench vat fitted with a wooden paddle, which may be pulled round by hand when necessary; in this way the skins are put under the liquor without danger of tearing or trapping them; in the case of grain splits the paddle is a great advantage over the pole.
The skins are allowed to rise two or three times, according to the condition required. For many kinds of alum leather the bate is omitted and the drench only is used.
In English practice the bran is mashed in the vat, in which the drenching is to take place, at a temperature of about 95° F. (35° C.); the skins are then put in and the drench well stirred or paddled until the whole is uniform, the temperature is then usually down to 85° to 83° F. (29·5° to 28·2° C.), at which temperature the fermentation begins. The temperature allowed varies with the state of the goods and the prevailing exterior temperature; in winter it may be higher than in summer, but never usually higher than 90° F. (32·2° C.). In some continental works the bran is mashed in boiling water, allowed to stand for some hours to cool, and only the bran-water is used; in this case the fermentation is different to that in which the bran is mashed at a lower temperature. In the first process the starch of the bran undergoes changes to dextrin and glucoses, which are then fermented by bacteria with the production of acids and gases (see Chapter X.), in the latter case the starch, which has been gelatinized is fermented by other organisms, which probably secrete diastatic enzymes. Less gas is given off in this fermentation than where the bran is mashed at 95° to 100° F. The temperature of fermentation is kept low, usually 25° C. (77° F.). Bran contains a sufficient amount of nitrogenous matter,151 to furnish the bacteria with nutriment, and until this is exhausted there is no fear of the skins being attacked.
There is also a “sweet” bran drench used on the continent, in which fermentation is not allowed to take place, the skins being paddled in an infusion of bran in water for a short time only, two to three hours at the most. In addition to the mechanical cleaning action, which the fresh bran has upon the skins, there is also a softening effect due to some constituent of the bran, which is at present not understood. One reason for this, is that the usual drench employed being an acid one, research has practically been confined to the latter.
Another method of preparing the bran drench, consists in allowing the required quantity of bran to soak in cold water for several hours; a sufficient quantity of hot water is then added to bring the temperature up to 50° C.; at this temperature the diastatic enzymes in the bran act rapidly on the starch and transform it into easily fermentable sugars. The temperature is allowed to fall to 34° to 35° C., and then a quantity of an old drench is added to start the fermentation. The liquor becomes acid, and in two days usually reaches its maximum acidity, when it is ready for use. The bran is strained off and only the milky liquor used. Such a drench is largely employed in the manufacture of chamois leather, the goods (sheep fleshes) being paddled in it for some hours previous to pressing. The French also use a similar process in the preparation of suede leather. The bran is strained off and only the sour partly fermented and fermenting liquor used. The straining off of the bran also saves working over the beam which would otherwise be required to get rid of the bran adhering to the skins.
It has recently been proposed to employ the bacillus bulgaricus, the organism which causes the fermentation of milk in the preparation of Bulgarian and Turkish “Yoghurt” (Bibliography 124) for drenching. Bacillus bulgaricus is capable of producing up to 2·5 per cent. of lactic acid, or about three times the quantity of that produced by ordinary lactic bacteria. Dr. Hugo Kühl (loc. cit.) proposes to cultivate the bacillus in the mother liquors, which are a by-product in the manufacture of milk sugar, and to use such liquor as a drench. Where skim milk or butter milk is to be had, this may also be used. A similar process is already in use in America, where the evaporated product is put upon the market under the name of Dermiforma (see pp. 189 and 194).
As a substitute for the drench, and in order to further remove lime after erodin, the late Franz Kathreiner employed a solution of anticalcium, 1 in 500 water, in which the skins remained overnight. Anticalcium is a mixture of sulphonic acids derived from cresols, and has antiseptic properties, so that its action is quite distinct from that of the bran drench. It is put on the market by J. Hauff, Feuerbach, near Stuttgart.
Drench Damage.—Drenching is a very important and useful process in leather manufacture, but, like other fermentive processes, requires great care and experience in its application. Although not so risky as puering, a good deal of serious damage may occur in the drench. The writer knows of no fuller discussion of all the various forms of drench damage than the articles of Eitner and others which appeared in “Der Gerber” during a long series of years (1882–1900). In these articles perhaps every known form of drench damage is described. Procter152 also gives a concise account of drenching. It will suffice to mention briefly the chief accidents to which the drench is liable; these are generally known as the “turning” of the drench.
1. Acid “turning.”—This usually occurs when the atmosphere is charged with electricity—during thunderstorms. It appears to be an extremely rapid form of butyric fermentation, which has not been fully investigated. The skins become swollen, transparent and tender, and, unless the fermentation is stopped, soon dissolve to a jelly. The only way to save the skins from destruction is to add salt to the drench; this reduces the swelling and, in fact, pickles the skin. I have pointed out elsewhere that this is really the origin of the modern pickling process, which is still known by the name of “rising.” A very acid drench will swell the skins almost the same as dilute sulphuric acid, and if they are then placed in a salt solution, or salt is added to the sour liquor, it is absorbed by the skins, and they are preserved almost as effectually as by the use of mineral acids.
2. Putrid “turning.”—This occurs under very similar atmospheric conditions to 1, and may be almost as rapid. Instead of becoming acid, the liquor turns slightly alkaline, frequently becoming bluish black, due to the presence of certain chromogenic bacteria. The goods fall as in a puer. Both the chromogenic and other bacteria present are peptonizing organisms, which obtain the upper hand owing to favourable conditions, and rapidly digest the skin unless the action is stopped. The best remedy is to remove the skins from the drench and pickle them without delay by the usual acid and salt process.
3. “Pinholey” drench.—This is the same as the Germans call “pikiren,” and is generally due to a too rapid evolution of gas both in the drench and in the capillary spaces of the skin itself. The gases form under the hyaline layer, and finally burst through in small holes. A damage very similar in appearance is also caused by colonies of gelatin liquefying bacteria developing on the grain. Each colony forms a small hole. The trouble is usually due to the setting of the drench at too high a temperature.
4. Spongy leather may be produced by leaving the goods too long in a sound drench, if they are allowed to rise too often. In this case experience alone can determine the proper duration of the drenching process.
A special case in which the hyaline layer is slightly attacked and its brilliance destroyed has been described by Eitner.153 A slimy film was formed on the surface of the drench and also on the grain of the skins. This film was found to consist of Bacillus megaterium (the potato bacillus), and where it was growing on the hyaline layer the latter was etched as it were, by the peptonizing enzyme secreted by the bacillus, so that when tanned the grain was covered with dull patches very much resembling “blast.”
Fermentations in the Drench.—Compared with the dung bate, or puer, the fermentation taking place in the bran drench is a simple one; as will be shown in detail in the next chapter, about half the weight of the bran consists of starch; this becomes hydrolysed by the enzymes in the bran (and in some cases secreted by the bacteria) into glucoses. According to the following equation:—
(C6H10O5)n + H2O = n C6H12O6
starch glucose
This expresses the change in its simplest form; in reality it is much more complex, various dextrins and sugars being formed simultaneously.
The sugars are then fermented by a variety of bacteria with formation of various organic acids, in accordance with the equation
C6H12O6 = 2 CH3CH(OH)COOH
which represents the formation of lactic acid.
In addition, formic and butyric acids are produced along with small quantities of other bodies, as will be shown. It is pretty certain that the butyric acid is formed by the further fermentation of the lactic acid and lactic salts in the liquid; at the same time CO2 and hydrogen are evolved as follows:—
2 C3H6O3 = C4H8O2 + 2 CO2 + H2
Fig. 27 is a drawing of the organisms seen in the drench, under a magnification of 1000 diameters. This was taken from a drench which had been set about 3 hours, and was just beginning to ferment; the bacteria form chains of varying length: the short chains have a slow motion in the liquor. As the fermentation proceeds, these form longer chains (Fig. 28), some of which are very beautiful objects, especially when viewed by dark ground illumination. The other organisms present are bacteria and bacilli from the puer, and at a later stage, as the nutrient material for the bran ferment becomes less, they begin to develop, first causing butyric and finally putrefactive fermentation.
A fairly pure culture may be obtained by inoculating a tube of bran infusion, previously sterilized, by means of a freshly made capillary pipette, which is pushed into the tube without removing the cotton-wool plug. The tube is placed in the incubator at 30° C., and the organisms causing the fermentation develop rapidly; as soon as the tube is observed to become cloudy, indicating the development of the bacteria, another tube is inoculated in the same manner, using a very minute quantity of liquid; this second “attenuation” is then carried on to five or six more tubes. In the fifth tube will usually be found a pure culture of the bran fermenting organisms, which I have called Bacterium furfuris. The bacteria (see Fig. 30) are mostly in the form of dumb-bells or pairs, each cell 0·75 µ × 1·25 µ: they vary slightly in size, some forming chains; those on the surface become surrounded by a kind of jelly, and form an iridescent pellicle on the surface of the liquid, the zoogloae form. When the nutriment of the liquid is exhausted or the acidity becomes too great,154 this film sinks to the bottom and the liquid above becomes comparatively clear. There is no spore formation, and, consequently, the culture soon dies unless inoculated into a fresh nutrient medium. The liquor, after the fermentation both in the drenches and tubes of pure cultivations, is always slightly ropy.
B. furfuris probably belongs to the group of coli bacteria, some of which ferment milk and other sugars with production of lactic acid and gases. This explains the ease with which the fermentation takes place, since the skins bring in immense numbers of bacteria from the bate, among which are the organisms described.
Eitner155 states that the action of the bran drench cannot be considered a chemical one, and that the chief action is a dynamical one, resulting from the gases generated in the liquid and in the skin. While I do not deny that the gas has such action—indeed I pointed out in 1893156 that the gas acted by floating and distending the skins, and so enabling them better to take up the acids—I maintain, however, that the chief action is a chemical one and due to the weak organic acids produced; this was confirmed experimentally. The researches on bran drenching commenced in 1887 by myself, and later in conjunction with Dr. W. H. Willcox, prove conclusively that the amount of acid produced, and especially the lactic acid, is sufficient to produce a very decided action on skin. In actual working we found from 1·07 to 2·34 grm. total acids per 1000 c.c., while in artificial cultures, with addition of calcium carbonate, inoculated with pure cultures of bacteria, obtained from drenches, we obtained from 4·53 to 11·44 grm. total acids per 1000 c.c. Now traces of lime in the skins act in just the same way as the calcium carbonate; they neutralize a portion of the acid produced by the bacteria, and so allow a greater amount of acid to be produced than if it accumulated in the liquid. By preparing solutions of the above acids of suitable strength and agitating skins in them, results were obtained in 1 12 hours equal to a drenching of 14 hours. The defect of such artificially prepared liquids in practice is due to the fact that they do not contain any particles of bran or flour; these take up dirt from the skin mechanically, and so produce a better-coloured leather. Where the skin has been bated in a perfectly clean bate, and then paddled in such an artificial drench, the leather is in every way equal to that prepared by means of the dung bate, followed by a bran drench.
By far the most important action for the tanner is the solution of the last traces of lime not dissolved by the bate, and this is the chief reason for its use, where colour is a desideratum, and where sweet liquors are used; for some kinds of leather where it is desirable not to lose pelt, the drench is used alone for this purpose. A dung bate, such as most light leather tanners use, is seldom run off more than once a week, and contains a considerable amount of dissolved lime; the amount in a freshly made-up puer wheel is from 0·1 grm. to 0·3 grm. lime (CaO) per litre. After the wheel has been in use for a week, it rises to 0·5 grm. to 0·8 grm. per litre. It will be readily seen that a liquid which already contains such considerable quantities of lime is not an especially good medium to use for its complete removal from the skins, hence drenching is necessary to get rid of the lime left in the skins.
Eitner states that the fermentation is alcoholic at the commencement, and then becomes an acid fermentation with generation of gas. In tan liquors there is no doubt that this is the course of the acetic fermentation, as has been shown by Andreasch,157 but in the usual type of bran drench the action is certainly a different one. The acetic acid, as far as we can ascertain, is produced directly from the dextrose, without the previous production of alcohol, since the presence of the latter is not shown by tests in any stage of the fermentation.158 No yeasts were observed159 and no alcohol was found in any stage of the fermentation.160 J. O’Sullivan remarks that although it has not been noticed it is possible that in this case the production of alcohol and its oxidation into acetic acid are simultaneous, but that there is no preliminary fermentation by yeasts followed by acetic fermentation by bacteria such as Eitner indicates is certain.
To sum up the conclusions arrived at from the experiments above quoted.
The starch of the bran or flour is first transformed into glucoses and dextrin by the action of an unorganized ferment called cerealin. The glucoses are then fermented by certain species of bacteria, Bacillus furfuris being perhaps the chief, with the formation of lactic, acetic, formic and butyric acids, and the evolution of carbon dioxide, hydrogen, nitrogen, and a small quantity of sulphuretted hydrogen.
The principal acid produced is lactic; the acetic acid is produced directly from the glucoses by the above-mentioned bacteria without any preliminary alcoholic fermentation by yeasts.
The mode of action of the drench on the skins may be summed up as follows:—
1. The solution of the last traces of lime which has not been removed by the bate, by the organic acids produced by the fermentation, and the subsequent swelling action of these on the skin fibres. The acids also dissolve a small amount of skin substance.
2. Simultaneously with (1) the distension and floating of the skins by gases produced by the fermentation, so enabling them better to take up the acids.
3. The mechanical absorption of dirt by the particles of bran or flour in the drench.
It is not proposed to enter into further detail, since the next chapter, consisting of the original papers of Dr. W. H. Willcox and the author, contains a pretty complete account of the analysis of the drench and theory of the drenching process.
1. Further Contribution on the Nature of Bran
Fermentation. By J. T. Wood & W. H. Willcox,
B.Sc.(Lond.).
(Reprinted from the Journal of the Society of
Chemical Industry, May 31, 1893. No. 5, vol. xii.
p. 422.)
The paper which one of us had the honour of reading before this Section on December 11, 1890,161 dealt chiefly with methods of bacteriological research, but especially in connection with the fermentation of bran as applied in the manufacture of light leathers. The object of the present paper is to give an account of further research into the nature of the fermentation and its products, the former communication being very incomplete. In further investigating the matter, we endeavoured—
1. To obtain a complete knowledge of the products of the actual fermentation as it takes place in practice.
2. To discover in what way the ferment acts, both on the materials fermenting and on the skins.
3. To examine in the same way the products of a pure cultivation of the bacterium causing the fermentation.
1. The Products of the Actual Fermentation.—These may be divided into three groups:—(1) gases; (2) volatile bodies; (3) non-volatile bodies. It was stated in the former paper that the ferment produced an inflammable gas along with considerable quantities of CO2, H2S, etc. The inflammable gas was thought by analogy from the researches of Tappeiner162 to be methane; it has, however, proved to be pure hydrogen. The absence of hydro-carbons was shown by the following method. Some of the gases were collected, the CO2 and SH2 removed by absorption with KOH, and the remaining gases exploded in a eudiometer tube with oxygen. The gases which remained after explosion did not diminish in volume after standing over KOH solution, showing absence of the paraffins and olefines.
For the purpose of analysis, about 1 12 litres of the gas were collected at a time in a large flask, fitted with a caoutchouc stopper and a funnel having an area of 28 sq. in. This was inverted, filled with the drench liquid, over the vat. The gas when collected was transferred immediately in the vat to a glass-stoppered bottle, sealed with a small quantity of the fermenting liquid and examined at once. Nine analyses have been made, most of them in duplicate. The following table, p. 248, gives the results of three of these duplicate analyses, which have been performed by Hempel’s method, the hydrogen being estimated by combustion in air over heated palladinised asbestos.
We find that the gases given off during the fermentation are practically the same, with or without skins.
Gases | A | B | C |
| CO2 and H2S | 21·9 | 25·2 | 42·4 |
| O2 | 1·0 | 2·1 | 3·6 |
| H2 | 53·1 | 46·7 | 28·2 |
| N2 | 24·0 | 26·0 | 25·8 |
| A is from a vat containing no skins, 1–2 days. | |||
| B from a vat containing skins, 2–3 days. | |||
| C from a vat containing skins, 3–4 days. | |||
The H2S is present only in small quantities from 1–2 per cent. Its presence was shown by aspirating the gases dissolved in 1 litre of drench through a dilute solution of lead acetate containing a few drops of acetic acid. The gases were liberated by heating the liquid, and at the same time aspirating air through it. The H2S is present both in the gases evolved from drenches which do not contain skins, and from those which do, though to a slightly greater extent in the latter. The amount of CO2 given off increases as the fermentation proceeds; the oxygen also increases, the nitrogen remaining practically constant. We consider that some of the nitrogen given off is that dissolved in the water, the oxygen being partly used up by the ferment in its earlier stages; the remainder of the nitrogen is probably produced from the decomposition of the nitrogenous bodies contained in the bran. It may be noted here that the bran fermentation under ordinary circumstances ceases on the fourth day, and sometimes earlier.
Frankland and Frew, in a paper on a pure fermentation of mannitol and dulcitol163 have shown the hydrogen and carbon di-oxide given off were produced by the decomposition of formic acid, the ferment producing formic acid and the latter immediately splitting up into an equal number of molecules of carbonic anhydride and hydrogen. We have every reason to believe, from experiments which will be included in the third section of the paper, that the source of the H and CO2 is the same in the fermentation we are considering.
2. Volatile Bodies.—These may be divided into (1) acids, and (2) amines. We had previously shown the absence of aldehyde by the rosaniline reaction, and of alcohol by Lieben’s iodoform test.
In the first experiment to determine the acids, 15,670 c.c. of the liquid from a normally fermenting vat was taken when the fermentation was at its height; this was submitted to distillation, and to the last portions distilled water added and 16,200 c.c. distilled over; the distillate was neutralised with sodium carbonate, and the whole was then evaporated in a porcelain dish, the residue dried first at 100° C., then over strong sulphuric acid, the weight of the sodium salts of the volatile acids thus obtained being 18·07 grm.
These salts were treated with 200 c.c. absolute alcohol and 20 c.c, strong H2SO4; heat was evolved, and there was a strong smell of ethyl acetate and butyrate.164 The mixture was allowed to stand for 24 hours and then distilled on the oil-bath, the temperature for a long time remaining at 81° C., finally rising to 96° C. 219 c.c. of distillate was obtained, and to this a saturated solution of common salt was added, but the esters of the volatile acids did not separate out. The whole was again redistilled with the same result.
Failing in this way to separate the acids in the form of their esters, the mixture of esters and alcohol was examined qualitatively; 75 c.c. was taken and saponified with 80 c.c. N/1 NaOH in a distilling flask, with inverted condenser, for half an hour, until all the fragrant smell of the esters had disappeared. 71 c.c. N/1 HCl was then added, and the apparatus connected with a condenser in the usual way. The distillate was acid. 800 c.c. was taken off, forming fraction 1. The remainder of the acid required to neutralise the sodium hydrate was added, and another 800 c.c. distilled off, forming fraction 2. Fraction 1 smelt strongly of butyric acid, fraction 2 of acetic acid. The fractions were then each boiled for half an hour with excess of barium carbonate; this was filtered off and washed, the filtrate evaporated to dryness, and dried at 130°.
A portion of barium salt of fraction 1 was heated with alcohol and sulphuric acid and gave the characteristic pineapple smell of ethyl butyrate.
The solution of the barium salts of fraction 2 was concentrated by evaporation, and to a portion excess of H2SO4 was added, the BaSO4 filtered off, and the filtrate carefully neutralised with ammonia and a portion added to a solution of neutral ferric chloride. It was coloured dark red, and a finely-divided precipitate of basic acetate of iron came down on boiling. A second portion of the concentrated solution was heated with sulphuric acid and gave off a strong smell of acetic acid. To a third portion silver nitrate was added, and a slight excess of acetic acid; a little silver was deposited in the form of a film, showing a trace of formic acid.
Having thus shown the presence of butyric, acetic, and formic acids in the drench, we proceeded to ascertain the quantities formed in a normal fermentation of bran without skins. For this purpose a drench was made in a clean vessel with 10 litres of distilled water and 200 grm. of bran mashed at a temperature of 38° C., and, after cooling to 33°, inoculated with bacteria from an actual drench which was fermenting vigorously; this was kept in the drench house so that the fermentation and general conditions might be exactly similar. In 48 hours all gases had ceased to be evolved, and the true fermentation was at an end. The liquid had an acid, not unpleasant smell, and was acid to litmus paper. The bran was strained off through muslin, washed with a little water, and well squeezed; the liquid measured 10 litres. Three litres of this were taken for the separation and estimation of the volatile acids, the remainder being set aside for examination and estimation of the non-volatile acids, etc. Two of the three litres were placed in a distilling flask with 5 grm. of pure CaCO3, and distilled down to one litre. The distillate was alkaline to litmus, and had a peculiar fishy smell; the remaining litre of drench was added, and the liquid taken down until the distillate ceased to be alkaline and only bad a faint smell. The alkaline distillate gave a yellow precipitate with Nessler’s solution, as well as the following reactions:—
| AgNO3 | A faint brown precipitate. |
| HgCl2 | A yellowish-white curdy precipitate. |
| PbA | A brownish-white precipitate. |
| CuSO4 | A dirty blue precipitate, which changed to a brownish turbidity on boiling. |
HCl was added to the distillate in slight excess and the liquid evaporated to a small bulk; to a portion chloroform and alcoholic potash were added, and the liquid heated; no smell of isocyanides was given off; the body is, therefore, not a primary amine. Phosphomolybdic acid gives no precipitate, therefore the body is not an alkaloid. From the above tests and its characteristic smell, we conclude that the body is trimethylamine. The platinum salt was formed by evaporating the above concentrated solution of trimethylammonium hydrochloride with excess of platinum chloride, a precipitate of the platinum salt insoluble in alcohol being formed. There was not, however, a sufficient quantity from the three litres to ascertain the molecular weight.
Proceeding with the estimation of the volatile acids, 100 c.c. N HCl being required to completely neutralise the CaCO3 used, 50 c.c. were first added and the distillation continued; the distillate was only very faintly acid. Four fractions were now distilled off, using respectively 10, 10, 10, and 20 c.c. N HCl.165
Fraction 1 was boiled with excess of barium carbonate, filtered, the BaCO3 washed with hot water, and the filtrate evaporated to dryness, and the Ba salts dried at 130° C. till the weight was constant; the salts were then decomposed with strong sulphuric acid, ignited, and the barium sulphate weighed.
Fractions 2, 3, and 4 were treated in an exactly similar way, the barium salts obtained and the weight and percentage of barium sulphate being shown in the following table:—
Fraction | Weight of Salts | Weight of BaSO4 | Per Cent BaSO4 | |
I. | 0·5585 | 0·4991 | 89·365 | |
II. | 0·5060 | 0·4618 | 91·265 | |
III. | 0·31675 | 0·29415 | 92·86 | |
IV. | 0·4475 | 0·4252 | 95·02 | |
Calculating the fractions 1 and 2 as mixtures of barium butyrate and acetate, and fractions 3 and 4 as mixtures of barium acetate and formate, we may summarise the results thus:—
Fraction | Weight of Salt | Ba Acetate | Ba Butyrate | Ba Formate | |
I. | 0·5585 | 0·4904 | 0·0681 | .. | |
II. | 0·5060 | 0·5030 | 0·0030 | .. | |
III. | 0·31675 | 0·2749 | .. | 0·04185 | |
IV. | 0·4475 | 0·2629 | .. | 0·18460 | |
| Total | 1·82875 | 1·5312 | 0·0711 | 0·22645 | |
or as free acids:—
Fraction | Acetic | Butyric | Formic | |
I. | 0·2308 | 0·0385 | .. | |
II. | 0·2367 | 0·0017 | .. | |
III. | 0·1293 | .. | 0·0170 | |
IV. | 0·1237 | .. | 0·0748 | |
| Total | 0·7205 | 0·0402 | 0·0918 | |
Proceeding now to (3) non-volatile bodies, we find these to consist of (1) acids, and (2) soluble carbohydrates. For examination, the remaining 4 litres of experimental drench were evaporated down to 1 litre and filtered; the residue (consisting of starchy matter) was well washed and the washings added to the filtrate, the whole placed in a large distilling-flask and distilled with continued addition of distilled water until the distillate was no longer acid (this required the addition and distillation of 4 litres of distilled water). A further deposit of solid matter rendering boiling dangerous, the liquid was again filtered and the residue washed free from acid. This residue was found to consist of nitrogenous organic matter and calcium phosphate, together with a trace of calcium oxalate, both being derived from the bran.
The clear liquid containing the non-volatile acids and other bodies was further concentrated and made up to 500 c.c.; it was dark brown in colour and was very acid to litmus; owing to a trace of flocculent matter, it was again filtered.
The presence of lactic acid was shown in the following manner: 10 c.c. of the liquid were placed in a small distilling-flask along with 2 c.c. strong H2SO4 and about 0·5 grm. potassium chromate in a little water. This was distilled and the vapours received in a test-tube surrounded by cold water; on adding magenta solution, decolorised by SO2, to the liquid in the test-tube, a red colour was produced by the aldehyde formed from the lactic acid; aldehyde was also recognised by its smell. We find this an exceedingly delicate test for lactic acid, and as far as we know it is quite new in this form.
For 10 c.c. of liquid to be examined, we find 2 c.c. strong H2SO4 and 1 grm. of potassium chromate to be the best proportions. Formic, acetic, propionic, butyric, valerianic, succinic, malic, tartaric, and citric acids do not give the reaction.
The liquid was now tested for succinic and malic acids. 25 c.c. was taken and decolorised by treatment with pure animal charcoal for half an hour; the liquid was then filtered, the charcoal washed free from acid, and the filtrate concentrated; ammonia was added in slight excess, the precipitate of calcium phosphate filtered off. To the filtrate CaCl2 was added in slight excess to remove the remainder of phosphates, the liquid filtered, and the filtrate cautiously neutralised with HCl; the addition of neutral ferric chloride to a portion gave no precipitate, showing the absence of succinic acid. To the remainder an equal volume of absolute alcohol was added and the liquid boiled; there was no precipitate. Twice the volume of absolute alcohol was then added and gave no precipitate. On the addition of four times the volume of absolute alcohol, a slight white precipitate came down, which evidently consisted of dextrin, from the manner in which it settled round the sides of the tube, and from its insolubility on adding HCl. We therefore conclude that there is no malic acid present, and that the only non-volatile acid produced is lactic acid.
The acidity was first determined by titrating 10 c.c. of the solution with N/10 sodium hydrate, using glazed litmus paper to determine the point of neutralisation. The 10 c.c. required 6·66 c.c. N/10 NaOH, equivalent to 0·666 of N lactic acid = 0·7481 grm. per litre of original drench.
In order to separate the lactic acid from the colouring matter and other bodies (dextrin and soluble starch) present, 100 c.c. of the concentrated liquid was decolorised with 10 grm. of animal charcoal, the mixture filtered and the charcoal well washed, the filtrate evaporated to dryness, and 15 drops of N/10 H2SO4 added to decompose any salts of lactic acid present; this was now extracted with ether, the ethereal extract of lactic acid placed in a distilling-flask, the ether distilled off, and the residue boiled with distilled water and pure calcium carbonate. The small amount of calcium sulphate present was removed by boiling the filtrate from the CaCO3 with barium carbonate, and filtering. The filtered liquid now consisted of a solution of calcium lactate; it was evaporated to dryness in a platinum dish, and the residue dried at 110° C.; it was then washed first with ether, then absolute alcohol.166
The insoluble residue was dried at 110° C. till the weight was constant.
The total weight of calcium salts obtained was 0·7661 grm. (from 45 litre of drench), of which 0·5493 grm. yielded on ignition 0·1398 grm. CaO = 25·45 per cent. Theory requires for calcium lactate (C3H5O3)2Ca = 25·69 per cent. CaO.
0·7661 grm. calcium lactate in the quantity used = 0·9576 grm. per litre = 0·7907 grm. lactic acid per litre. The difference between this and the preceding amount of lactic acid found by titration, viz. 0·0426 grm., is probably accounted for by the presence of a small quantity of salts of lactic acid in the drench.
The second part of the subject which we proposed to consider was “In what way does the ferment act on the bran and on the skins?” The average composition of bran is shown in the following table:—
Per Cent. | |
| Water. | 14 |
| Fibrin, etc. | 15 |
| Starch | 44 |
| Fat | 4 |
| Lignose and cellulose | 17 |
| Ash | 6 |
It will be seen, as stated in the former communication,167 that the starch must be the principal body acted upon; but the cellulose is also an important constituent, and before going further it was necessary to ascertain if it took part in the fermentation. For this purpose some pure cellulose was prepared from cotton wool in the usual way, and small portions placed in tubes containing yeast-water168 as a nutrient material. These were sterilised by steaming; two tubes were inoculated from a pure cultivation of the bacteria obtained in 1889, two were inoculated from an actual drench, and three left uninoculated; all of them were placed on the incubator at a temperature of 30°–33°. On the second day the inoculated tubes were cloudy, but no gas was given off, nor was any acid formed; in 10 days the cellulose had not disappeared, nor on examination with the microscope could any action be detected. The experiment was repeated with peptone as a nutrient medium, but with the same result. The conclusion is that the bacterium does not attack the cellulose, which thus takes no part in the fermentation. The starch and nitrogenous bodies of the bran are therefore the only bodies acted upon by the bacteria in this fermentation.
From the fact that bran drenches ferment in the same way when mashed at all temperatures from 20° C. to 40° C., and that in all cases the starch is decomposed, it was supposed that the ferment was capable of attacking the starch in its undissolved condition. To ascertain if this were so, it was necessary to use pure cultivations in the laboratory.
The usual methods employed had thrown no light on this part of the subject, as in order to sterilise the solutions they had been repeatedly boiled, and were thus not comparable with the fermentation as it takes place in the works.
In order to get rid of this difficulty, the starch was sterilised in a dry condition in the hot-air oven, by heating for several hours on successive days to 110° C.
This sterile starch was mixed with sterilised water in tubes plugged with sterile cotton-wool. Eight tubes were taken, as follows:—
1. Sterile starch and water.
2. Sterile starch and water inoculated pure culture.
3. Sterile starch and yeast-water inoculated pure culture.
4. Sterile starch and asparagin inoculated pure culture.
5. Yeast-water alone inoculated pure culture.
6. Dextrin169 and yeast-water inoculated pure culture.
7. Soluble starch170 and yeast-water inoculated pure culture.
8. Starch mucilage and yeast-water inoculated pure culture.
These were allowed to stand on the incubator at 33°–35°C., and examined for acid each day by the method described in the previous communication for starch testing. They all remained neutral, although the bacteria developed in all but No. 1. These experiments were repeated several times, with the same result in every case. They show that this ferment is unable to act on starch either in its insoluble or soluble condition, alone or in the presence of nitrogenous bodies.
Now it has been known for a considerable period that bran contains an unorganised ferment called cerealin,171 which is capable of changing starch into dextrin and other carbohydrates; but the information to be obtained about it was very meagre, and as it appeared that this body might play an important part in the fermentation, we proceeded to prepare some pure cerealin and to ascertain its action on pure starch. The cerealin was prepared by taking a kilo of bran and extracting it with 2 litres of distilled water at 30° C; the extract was filtered clear and 2 litres of strong spirits of wine containing 90 per cent. alcohol was added, when a flocculent precipitate separated, which was washed on a filter with alcohol, dehydrated with absolute alcohol, and dried over H2SO4. The cerealin thus prepared is an amorphous substance not quite white, difficultly soluble in water, though we think this is due to its having been coagulated, and that it might be prepared in some other way which would show it to be more soluble than that which you now see.
To show its action on starch we took 10 grm. of pure starch in 200 c.c. water at 40° C., and placed equal quantities in two flasks; to No. 1 about 0·1 grm. of the cerealin was added; No. 2 was left blank. These were kept at 40° C. for 10 hours, the clear liquid filtered off, and examined with Fehling’s solution. No. 1 reduced it strongly, showing that glucoses were present in considerable quantity. No. 2 had no effect whatever.
Addition of alcohol to No. 1 gave a white precipitate; this was thrown on to a filter, washed with alcohol, and dried at 100°, again dissolved in water, and inverted by boiling the solution with 120 part of strong sulphuric acid and then neutralising with sodium hydrate. The resulting solution reduced Fehling’s solution, showing that the body was dextrin. Strong bran infusion (which of course contains cerealin) acting on a thick starch mucilage, liquefies it, and forms glucoses and dextrin.
We have thus shown that the cerealin produces glucoses as well as dextrin, both from solid starch and from starch mucilage. This is most important, as it has been previously shown by one of us that the ferment attacks glucose very easily.
The drenches were now examined in order to ascertain the presence of glucose and dextrin; in the former communication it was stated that these were absent. We find, however, that by concentrating the liquid, that both are present in the early stages. Samples were taken one hour after mashing, and at 3, 6, 12, and 18 hours, while the drench was working.
These were evaporated to 15 bulk, filtered, and divided into two portions, one of which was examined with Fehling’s solution; to the other alcohol was added; the white precipitate after addition of alcohol was filtered off, washed with alcohol, dried at 100° C., redissolved in water, and boiled with sulphuric acid to invert it, then examined with Fehling’s solution.
The following results were obtained:—
Hours | 1 | 3 | 6 | 12 | 18 |
| Glucoses or sugars reducing Fehling’s solution directly | Present in considerable quantity | Present in small quantity | Absent | Absent | Absent |
| Dextrins, Fehling’s solution reduced after inversion | Present in considerable quantity | Present | Traces | Faint traces | Absent |
| Soluble starch | Absent | Absent | Absent | Absent | Absent |
In a bran infusion kept from fermenting by a little ether or chloroform, the formation of glucose and dextrin goes on continuously, the glucose increasing in quantity; the action is, however, much slower than in the case of diastase; at the end of 12 hours, at a temperature of 40° C., about half the starch is transformed.
It appears from this and a number of other experiments that glucoses and dextrin are formed by the cerealin, the former only being decomposed by the bacteria almost as fast as it is produced, for after three hours no glucose is found in the drenches.
We have thus shown that the acids and gases are produced from the starch contained in the bran, the starch being first changed into glucoses by the action of an unorganised ferment or ferments; and that the glucoses are decomposed by a specific organism, the nitrogenous material in the bran serving for its nutriment; that the action is the same with or without skins although there appears to be a little more H2S gas given off from drenches containing skins, than from those containing none.
The ferment has no direct action on the skins. This may be shown by taking a piece of limed skin, in which a considerable portion of the lime exists as carbonate, and submitting it to the action of the ferment; in this case the action goes on much longer than in the drenches, being complete in about 15 days, but the skin may be left in the resulting liquid for three months without undergoing further change than solution of the lime, provided that suitable means be taken to exclude moulds, which, by destroying the organic salts and acids produced, enable putrefactive fermentation to begin.
It has been thought that the bran itself exercised some peculiar action on the skin, and possibly this may be so to a slight extent,172 as the sweet bran drench is occasionally used on the continent, but if skins are placed in a mixture of bran and water (in the proportions for drenching) which is prevented from working by the addition of a minute quantity of HgCl2 110000, such a drench has no action on them, and when tanned they are harsh and hard, similar experiments have been made on a smaller scale, using ether and chloroform to prevent fermentation, with the same results.
In order to show whether the acids alone were the cause of the action on the skins, an artificial drench was made up of the following composition—
0·5 grm. per 1000 glacial acetic acid.
1·0 " " " lactic acid sp. gr. 1·210.
In this skins were worked intermittently for 1 12 –2 hours, and it was found that in this time they were in a similar condition to skins which had been in a drench from 12–16 hours. They were afterwards tanned, and found to be good leather, and in every way equal to similar skins which had been “drenched.” A number of experiments have been tried with sulphuric and hydrochloric acids in order to ascertain if these had a similar action, but the results have not been satisfactory.
With regard to the third portion of the research, viz. the products of a pure cultivation of the bacteria, we have obtained a good number of results; but as the description of the experiments is of a greater length than we anticipated, and as there is still some work to do in verifying them, we are obliged to leave this portion for another paper, which we hope to have ready by the next session.
In conclusion, we may summarise the results obtained up to the present in the fermentation of bran by the organism we have used; remarking that there may be other organisms capable of fermenting bran in a somewhat similar manner.
1. It has been shown that the fermentation investigated is due to a specific organism, of which we find no account, and which, pending further experiments, we have therefore provisionally named Bacterium furfuris.
2. That the starch and nitrogenous bodies in the bran, alone take part in the fermentation, the starch being first transformed into glucoses and dextrin by the action of an unorganised ferment or ferments; the glucoses and nitrogenous bodies only, being decomposed by the bacteria, with the formation of formic, acetic, butyric, and lactic acids, and the simultaneous evolution of hydrogen, carbon dioxide, nitrogen, and a small quantity of sulphuretted hydrogen. The following table shows the quantities found in an experimental drench per 1000 c.c.
Grm. | |
| Formic acid | 0·0306 |
| Acetic acid | 0·2402 |
| Butyric acid | 0·0134 |
| Lactic acid | 0·7907 |
──── | |
| Total | 1·0749 |
──── |
We find in actual work that the quantity of acid produced varies from 1 to 3 grm. per litre.
3. That if these acids are applied to the skins in the same proportions as they occur in the drench, the action on them is the same, and much quicker than an ordinary drench.
4. That the gas therefore, has no action on the skins per se, with the exception of floating and distending them, and so enabling them better to take up the acids.
We are indebted to Mr. H. R. Procter, of the Yorkshire College, Leeds, and to Dr. Percy F. Frankland for valuable suggestions in carrying out some of the work.
II. On a Pure Cultivation of a Bacillus Fermenting
Bran Infusions. By J. T. Wood and
W. H. Willcox, B.Sc.(Lond.).
Reprinted from the Journal of the Society
of Chemical Industry, 30th June, 1897, No. 6,
vol. xvi. p. 510.
Isolation of Pure Culture.—In our previous communication on bran fermentation as applied in the manufacture of light leathers,173 we gave an account of the actual fermentation and its products, together with the mode of action on the bran and on the skins for which this fermentation is used, reserving to the present paper an account of the products of a pure cultivation of the bacteria causing the fermentation.
The cultivation used in the first experiments for this purpose was one isolated in 1889, and used in the cellulose and starch experiments described in the above-mentioned paper.
This cultivation had not been obtained from a single colony from gelatin, and in order to make quite sure that the cultures used were pure, it was decided to make another attempt to isolate the bacillus by plate cultivation. Previous attempts to do this had failed, bate organisms and gelatin liquefying bacilli, developing in such numbers that the plates were spoiled before the organism, which caused the fermentation, had time to develop; beside which the organisms, as obtained direct from the drenches, grew with difficulty in the ordinary nutrient gelatin. A special gelatin was therefore prepared of the following composition:—
| Gelatin | 100 grm. | |
| Glucose | 30 " | |
| Salt solution | 200 c.c. | |
| Water | 800 " |
Plates of this gelatin in Petri dishes were prepared from the previously used supposed pure cultures which had been preserved in sealed tubes. These were found to be dead. A modification of the method previously described by one of us175 was adopted.
A solution of nutrient glucose was inoculated from a working drench, and as soon as the liquid was observed to become cloudy, a tube of the solid glucose gelatin was inoculated from it by plunging in a platinum needle. In two days the bacteria developed along the needle track. Fig. 32 shows the appearance of the tube four days after inoculation, a bubble of gas being formed in the solid gelatin. On the following day, the tube was broken, and from the portion where gas was given off most vigorously other tubes of solid and liquid media were inoculated. Acid was quickly formed in the nutrient glucose solutions. In the gelatin tubes, the bacteria developed well in the depth. The now purified culture was passed through three more glucose gelatin tubes, each time also a glucose tube being inoculated. From the last of these tubes a very minute quantity was taken 12 hours after inoculation on the point of a platinum needle, and a streak culture made on glucose gelatin. In 24 hours a growth could be seen on the surface of the gelatin in the form of minute dots perfectly separated one from another.
Organisms causing Bran Fermentation.
Pure Cultures.
From one of these dots a tube was inoculated and from this several plate cultivations were made. The colonies which developed on these plates were of two kinds, the majority being round, yellowish and of small size, a smaller number spreading out on the surface of the gelatin and slightly iridescent. These surface expansion colonies when examined with a low power appear like a milky drop, with very fine granular contents, the whole surrounded by wavy lines which follow exactly the irregular contour of the expansion. The small round colonies growing in the depth occur in the proportion of about 3 to 1 of the surface expansion colonies. The microscopic appearance of the bacteria composing the two kinds of colonies, is almost exactly similar, they are extremely small and regular in size, 0·75 µ × 0·5 µ to 0·7 µ × 1 µ. When spread upon a slide, they are not readily miscible with water, and appear greasy. Both colonies inoculated into glucose tubes produced acid. The existence of these two organisms was confirmed in the following way:—A glucose tube was inoculated from a drench in active fermentation; as soon as the liquid became cloudy, a second tube was inoculated from it by means of a platinum needle; from this tube the fermentation was carried through two more tubes; a plate cultivation was made from the last tube 10 hours after inoculation. Again, the two kinds of colonies developed exactly similar in every respect to those obtained from the streak cultures.
It seems probable from these results, and also from a comparison of the fermentations made with the organisms from an actual drench,176 and from purified cultures with those from a single organism, which are described in the present paper, that the action in the drenches is a symbiotic one in which two or more organisms take part.
The Fermentations.—During the time occupied by the isolation of pure cultures of the bacteria, two fermentations were conducted with the supposed pure cultures. These fermentations (or rather the second of them, for the first was unfortunately lost through the breakage of a bottle) may prove of considerable interest as throwing some light on the symbiotic action of the two organisms.
The first fermentation with pure cultures of the bacillus α (B. furfuris) obtained from a single colony in glucose gelatin, was inoculated on September 16, 1894, the composition of the fermenting liquid being—
| Glucose | 27 | grm. |
| Peptone | 1·4 | " |
| Salt solution | 140 | c.c. |
| Water | 860 | " |
| Pure calcium carbonate | 10 | grm. |
This was contained in a narrow-necked litre flask fitted with a rubber stopper, and narrow delivery tube dipping under mercury, and sterilised with all the usual precautions. The fermentation began on the second day, reached its height from the 6th–8th day, and continued for 39 days, when gas ceased to come off. The examination of the gases will be described later on. When the fermentation was over, the liquid was brought to boiling temperature. It was then examined for the volatile acids in exactly the same manner as we described in our previous paper.
140 c.c. normal HCl was added and distillation commenced; the distillate was acid. The distillation was continued until the distillate ceased to be acid, forming fraction I. Three more fractions were now distilled off using respectively 10, 20, and 17 c.c. N1HCl.
The fractions were boiled with excess of BaCO3 filtered, the BaCO3 washed with hot water, the filtrate evaporated to dryness, and the barium salts dried at 130° C. till the weight was constant.177 The salts were then decomposed with strong H2SO4, ignited, and the barium sulphate weighed. The following is a tabulated statement of the results:—
Fraction | Weight of Salts | Weight of BaSO4 | Per Cent. of BaSO4 | |
I. | 1·2420 | 1·0915 | 87·88 | |
II. | 0·9915 | 0·9170 | 92·49 | |
III. | 1·2155 | 1·1980 | 98·56 | |
IV. | 0·6350 | 0·6230 | 98·11 | |
Calculating fraction I. as a mixture of barium acetate and butyrate, and fractions II., III., and IV. as mixtures of barium acetate and formate,178 we get:—
Fraction | Weight of Salts | Ba Butyrate | Ba Acetate | Ba Formate | |
I. | 1·2420 | 0·2630 | 0·9790 | .. | |
II. | 0·99151 | .. | 0·89330 | 0·0982 | |
III. | 0·2155 | .. | 0·44000 | 0·7755 | |
IV. | 0·6350 | .. | 0·2552 | 0·3798 | |
| Totals | 4·0840 | 0·2630 | 2·5675 | 1·2535 | |
Calculating the barium salts into their respective acids we get:—
Fraction | Formic Acid | Acetic Acid | Butyric Acid | |
I. | .. | 0·46070 | 0·1488 | |
II. | 0·03980 | 0·42040 | .. | |
III. | 0·3143 | 0·20710 | .. | |
IV. | 0·1539 | 0·1201 | .. | |
| Totals | 0·5080 | 1·2083 | 0·1488 | |
The total volatile acids produced amounting to 1·8651 grm.
The residual liquid containing the non-volatile acids was submitted to the test for lactic acid previously used,179 and it was found to be present.
The method employed for estimating lactic acid in our previous communication proving somewhat difficult, we endeavoured to improve it by extracting the concentrated solution of the non-volatile acids on prepared pumice stone with ether in a paper thimble contained in a Soxhlet fat-extraction apparatus. After repeated trials we found that this method did not give accurate results. The solution was therefore titrated with 110 N sodium hydrate, using glazed litmus paper to determine the point of neutralisation. The acidity found corresponded to 2·438 grm. of lactic acid per 1000 c.c. of the fermented liquid.
We have done several other fermentations with this organism and find the same acids produced and the same gases evolved, the results just given being fully confirmed. At the same time the amount of the acids produced and their proportions vary, that is to say, the quantity of acid from a given fermentation cannot be predicted with absolute accuracy, although the conditions under which we carried out the experiments were made as like as possible.
We give the total acids from four fermentations to show the amount of variation. I. is a symbiotic fermentation caused by organisms α and β; the remainder are fermentations by α alone.
Fermentation | I | II | III180 | IV | ||||
| Total vol. acids, grm. per 1000 c.c. | 2 | ·4968 | 1 | ·8651 | 0 | ·9738 | 1 | ·5636 |
| Mean percentage BaSO4 from Ba salts of vol. acids | 89 | ·17 | 91 | ·76 | 95 | ·4 | 93 | ·6 |
| Lactic acid | 8 | ·9500 | 2 | ·4380 | 1 | ·4737 | 2 | ·9700 |
Comparison of Acids from Fermentation II. and III.180
— | Lactic | Formic | Acetic | Butyric | ||
II. | 2·4380 | 0·5080 | 1·2083 | 0·1488 | ||
III. | 1·4737 | 0·3914 | 0·5593 | 0·0231 | ||
Fermentation III., 2000 c.c.
Fraction | Weight of Salts | Weight of BaSO4 | Per Cent. of BaSO4 | |
I. | 0·7260 | 0·6535 | 90·01 | |
II. | 0·8150 | 0·7410 | 90·92 | |
III. | 2·1525 | 2·1342 | 99·15 | |
IV. | 0·3695 | 0·3627 | 98·16 | |
V. | 0·3530 | 0·3500 | 99·15 | |
Calculation of Barium Salts as Barium Butyrate, Acetate, and Formate.
— | Ba Butyrate | Ba Acetate | Ba Formate | |
I. | 0·0597 | 0·6663 | .. | |
II. | 0·0223 | 0·7927 | .. | |
III. | .. | 0·6662 | 1·4863 | |
IV. | .. | 0·1469 | 0·2226 | |
V. | .. | 0·1070 | 0·2440 | |
| Totals | 0·0820 | 2·3791 | 1·9529 | |
Equivalent to 0·0463 grm. butyric acid.
" 1·1186 " acetic "
" 0·7828 " formic "
or one-half of these quantities per litre of the fermented liquid.
Note.—Mr. Adrian J. Brown, of Burton-on-Trent, has been kind enough to examine a sample of the glucose used in the above fermentations, and found the rotary power to be equivalent to 95·6 per cent. pure dextrose. The zinc salt of the lactic acid produced had no rotary power.
The Gases.—In dealing with the gases evolved, we first compare those given off in the fermentation of glucose with that of bran under exactly similar conditions. The fermentation was conducted in open vessels as before described,181 and the gases were collected and examined in the same way.
Mean of three Analyses.
— | Bran and Skins | Glucose and Skins |
| CO2 | 25·2 | 24·5 |
| O2 | 2·1 | 1·5 |
| H2 | 46·7 | 49·8 |
| N2 | 26·0 | 24·2 |
The composition of the gases is thus almost exactly similar, and, we think, fully proves our previous conclusions as to the change of the starch of the bran into glucoses by means of an unorganised ferment (cerealin).
In the closed fermentations we had previously collected only small quantities of gas over mercury, owing to the difficulty of continuously collecting large quantities which came off during the night.
In the fermentation of September 16, 1894, we collected the whole of the gas given off, taking samples every day over mercury, the gas coming off at night being collected over warm water. Of course this method does not give the total amount of gas evolved with absolute accuracy, but the exact composition of the gases was known from day to day, and the amount of CO2 absorbed by the water could be calculated with moderate accuracy.
The fermentation was conducted in a narrow-necked litre flask fitted with a narrow delivery tube dipping under mercury, and sterilised with all the usual precautions. The temperature was maintained at 25°–30°, gas was evolved for 39 days, when it ceased to come off, the total amount collected being 3435 c.c. One-half of this quantity, however, came off in seven days. About 300 c.c. of CO2 was absorbed by the water during the whole period. The diagram (Fig. 33) shows the manner of evolution of the gases, the ordinates representing volume of gas and the abscissæ lapse of time after inoculation. The following table shows the composition of the gas at different stages of the fermentation. (The fermentation (II.) is the one of which the chemical analysis has been previously given, page 273):—
Composition of Gases evolved in Fermentation of 1000 c.c.
Glucose with pure Ferment. September 16, 1894.
— | Day | Mean | |||||
5 | 9 | 12 | 19 | 26–31 | 35–39 | ||
| CO2 | 53·7 | 56·4 | 44·3 | 52·6 | 55·6 | 43·2 | 49·9 |
| O2 | 1·8 | 0·5 | 1·7 | 1·9 | 0·8 | 2·8 | 1·8 |
| H2 | 35·8 | 34·6 | 41·2 | 30·3 | 34·7 | 31·8 | 34·8 |
| N2 | 8·7 | 8·5 | 12·8 | 15·2 | 8·9 | 22·2 | 13·5 |
The total quantity of CO2 actually collected = 1563 c.c. = 3·090 grm.; the amount of CO2 due to decomposition of the CaCO3 by the acids produced was found to be 667 c.c. = (1·3189 grm). The vol. of hydrogen collected was 1086 c.c. = 0·973 grm.
In a second fermentation (III.) we endeavoured to ascertain the exact amount of CO2 evolved, as in the previous fermentation this had not been done. It was therefore decided to absorb the CO2 by means of potash.
The fermentation in this case was conducted in a narrow-necked flask of 2000 c.c. capacity, connected by means of a narrow glass tube with two potash bulbs containing strong caustic potash, and furnished with a delivery tube dipping under water; the whole apparatus stood upon an iron plate, and was maintained at a temperature of 25°–30° in the same manner as the previous fermentation. The gases were evolved for 21 days—a considerably shorter period than the 1000 c.c. fermentation; but resembling it in that one-half the gas was evolved in eight days. The diagram shows the curve as in the previous fermentation, which it resembles for the first 14 days, afterwards however stopping suddenly. When the fermentation was at an end the flask and contents were heated to boiling point, at the same time a current of air free from CO2 was drawn through it, and the CO2 given off being collected in potash bulbs as in the fermentation. Unfortunately the estimation of the CO2 was rendered valueless owing to an accident.
The table shows the composition of the gases other than CO2 evolved in this second fermentation.
Gases from Fermentation of 2000 c.c. (excluding CO2)
Fermentation III.
— | Days | Mean | |||||
2–4 | 4–5 | 5–6 | 11 | 14–15 | 16–17 | ||
| O2 | 3·4 | 2·2 | 1·5 | 0·42 | 0·9 | 2·4 | 1·48 |
| H2 | 81·3 | 83·3 | 82·4 | 79·0 | 71·7 | 72·2 | 77·72 |
| N2 | 15·3 | 14·5 | 16·1 | 20·5 | 27·4 | 25·4 | 20·78 |
The gas from days 18–21 was unfortunately mixed with air. On comparing the mean composition of gases other than CO2 collected from both fermentations, we get the following result:—
— | Fermentation, 1000 c.c. | Fermentation, 2000 c.c. |
| O2 | 3·57 | 1·48 |
| H2 | 69·4 | 77·72 |
| N2 | 27·0 | 20·78 |
If now the O and part of the N in the proportion to form air be taken away, the composition of the gases from the two fermentations is found to be almost exactly similar:—
— | Fermentation, 1000 c.c. | Fermentation, 2000 c.c. |
| H2 | 84·4 | 83·9 |
| N2 | 15·6 | 16·1 |
100.0 | 100.0 |
The gases from a third fermentation were almost exactly similar in composition, but the total volume was not measured.
A remarkable fact in this fermentation is the evolution of free N, which seems to be rare, except in the case of putrefactive organisms, as in the vast number of fermentative decompositions due to bacteria, almost the only gases found are carbonic anhydride, hydrogen, H2S, and marsh gas.
Gayon182 in 1875, in an investigation on the putrefaction of eggs, collected the gas given off from large ostrich eggs, and found in it 29 per cent. of nitrogen; he adds, however, that its presence may be due to the accumulation of a certain quantity of air in the air-bubble before putrefaction.
Béchamp183 found that yeast cells under suitable conditions, but sugar being withheld, produced pure nitrogen along with leucin, tyrosin, a soluble albuminous substance coagulable by heat, an enzyme, a peculiar gummy substance, phosphates and acetic acid, alcohol and CO2. These are almost the only instances where observers of repute have been convinced of the evolution of free N by bacteria. We find since the above work was carried out that Immendorf184 has found certain bacteria in dung which form ammonium nitrate, and this body, as is known, splits up at a comparatively low temperature into nitrogen and water.
From the bacteriological as well as the chemical results, it is now evident that the fermentation as it takes place in practice is a symbiotic one in which two organisms play the most important part, and very probably cause the entire fermentation. This is shown by comparing the acids produced by the fermentation in the works with those produced by a mixture of the organisms α and β, the relative amounts being very close, while in all the fermentations with α alone a much less proportion of lactic acid is produced, as the following table shows:—
— | Fermentation in Works | Fermentation α and β | Fermentation α (II.) | Fermentation α (III.) |
| Formic acid | 2·8 | 0·8 | 11·8 | 16·0 |
| Acetic " | 22·5 | 16·4 | 27·9 | 22·7 |
| Butyric " | 1·2 | 4·5 | 3·5 | 0·9 |
| Lactic " | 73·5 | 78·3 | 56·7 | 60·2 |
Note.—Ruge (Sitzungsber. d. Wien. Akad. d. Wiss, Vol. xliv., 1862, 734) found that the gases of the large intestine contained 57·8 per cent. of N, and Gamgee remarks “in part a diffusate from the blood, but is certainly in part derived from the bacterial decomposition of proteids.” (Gamgee, Phys. Chem. Vol. ii., p. 467.)
The acetic acid, as far as we can ascertain, is produced directly from dextrose without the previous production of alcohol, since the presence of the latter is not shown by its tests at any stage of the fermentation. We have also ascertained that the organism is without action on dilute solutions of alcohol, in yeast water, no acid being produced.
We are indebted to Mr. H. S. Shrewsbury for the analysis of some of the gases and volatile acids, and also for the preparation of the diagrams. In conclusion we may state that the investigation of this fermentation in the tannery has been the means of pointing the way to a still more complicated process, viz., “bating.” It may even be possible in the future to place these processes on somewhat the same footing as the accurately understood fermentations in the brewing industry although the difficulties in the way are much greater.
The following short bibliography of works on tanning and the bacteriology of leather manufacture, does not profess to be complete, but is given in the hope that it may prove useful. It includes most of the works consulted or referred to in this book. For particulars of some of the earlier works (Nos. 1, 2, 4, 5, 6, 7, and 8), I am indebted to Dr. E. Stiasny, of Leeds University.
Nos. 1–10 are old works on Tanning generally.
Nos. 11–57 books and papers referring chiefly to bating and the bacteriology of Leather Manufacture.
Nos. 58–78 deal with putrefaction.
Nos. 79–133 are general works consulted, in order of date.
1. La Tannerie et la preparation des Cuirs. (MS.) Desbillettes. 1708.
2. L’art du tanneur. De la Lande. 1764.
3. The art of tanning and currying leather, with an account of all the different processes made use of in Europe and Asia for dyeing leather red and yellow, collected and published at the expense of the Dublin Society. To which are added Mr. Philipo’s method of dyeing the Turkey leather as approved of by the Society for the Encouragement of Arts, etc., and for which he had a reward of £100 and their gold medal for the secret. Also the new method of tanning invented by the late David Macbride, M.D., London, reprinted for J. Nourse in the Strand, Bookseller to His Majesty. 1780.
4. Lohgerberei, Ignatz Bautsch. Dresden, 1793.
5. Ueber die Bearbeitung der Tierhäute zu allen Gattungen von Leder. P. J. Kasteleyn. German translation from the Dutch. Leipzig, 1797.
6. Chemisch-technologische Grundsätze der gesammten Ledergerberei. 2 vols. S. F. Hermbstädt. Berlin, 1805 and 1807.
7. Dictionary of Chemistry and Mineralogy. 2 vols. A. and C. R. Aikin, 1807.
8. Hand-Encyclopädie für das Gerben, Zurichten, etc., des Leders, L. F. Kummer, Berlin, 1830.
9. Handbuch der Gesammten Lohgerberei. Vom Dr. Ch. H. Schmidt. Weimar, 1847.
10. Lehrbuch der Sohlledergerberei. Dr. G. W. Bichon, Berlin.
11. “Das Beizen der Glacé-Felle” in Handbuch der Weissgerberei. Anton Brüggemann. Quedlinburg and Leipzig, 1857. p. 21.
12. Erfahrungen auf dem Gebiete der Gerberei. J. C. H. Lietzmann, Berlin, 1862.
13. “Das Behandeln in der Kleienbeize” in Handbuch der Weissgerberei. Dr. W. F. Gintl. Weimar, 1873. p. 51.
14. Mistbeizen. Der Gerber, 1884. p. 197.
15. The Manufacture of Leather. Davis, Philadelphia, 1885. p. 335, etc.
16. Traité pratique de la Fabrication des cuirs et du Travail des peaux. Villon, 1889. p. 407.
17. Die Englische Methode für die Chevrettengerbung. Beizverfahren, Der Gerber, Bd. XV. 1889. p. 267.
18. Methods of Bacteriological Research, with some account of Bran Fermentation, by J. T. Wood. Jour. Soc. Chem. Ind. IX. 1890. p. 27.
19. The Theory and Practice of Tanning. W. J. Salomon. Tech. Quarterly, 1892. 5 (1 and 2), 81–88.
20. Further contribution on the Nature of Bran Fermentation, by J. T. Wood, and W. H. Willcox, B.Sc., London. Jour. Soc. Chem. Ind. XII. 1893. p. 422.
21. The Chemistry of the Grainer Pit, by T. Palmer and P. G. Sandford. J. Anal. and Appl. Chem., 1893, 7, 87–95.
22. Fermentation in the Leather Industry, by J. T. Wood. Jour. Soc. Chem. Ind. XIII. 1894. p. 218.
23. The Bacteria of Stable Manure and their Action. Dr. E. Herfeldt, Bonn. Jour. Soc. Chem. Ind. XIV. 1895, pp. 449–452. Translated with foreword by J. T. Wood. (This paper contains a Bibliography of 21 items.)
24. Uber die Beziehung der Bakteriologie zur Gerberei. Dr. F. H. Haenlein. Cent. Bl. f. Bakt. II. 1895. p. 26.
25. Die im Miste vorkommenden Bacterien und deren physiologische Rolle bei der Zerzetzung desselben. S. A. Severin. Central Bl. f. Bakt. (II), Vol. I., 1895, pp. 97, 160, 799. Vol. III. pp. 628, 706.
26. Gährungserscheinungen in Gerbbrühen. Fr. Andreasch. Der Gerber, 1895–7.
27. W. Schmitz Dumont. The Sweating Process in the Tannery. Ding. Polyt. Jour. 1896, 300, 139–144. J.S.C.I. 1896, p. 461.
28. On a pure cultivation of a Bacillus fermenting Bran Infusions, by J. T. Wood and W. H. Willcox, B.Sc., London. Jour. Soc. Chem. Ind. XVI. 1897. p. 510.
29. The Rationale of Bating, by J. T. Wood. Leather Industries. September, 1898.
30. Notes on the constitution and mode of action of the Dung Bate in leather manufacture. J. T. Wood, Jour. Soc. Chem. Ind. XVII. 1898. p. 1010.
31. “Schwitzen” in Gerberei Chemie, Sammlung von Aufsätzen Professors Dr. Julius von Schroder, 1886–1895, published 1898.
32. Beizen und neue Beizmittel, W. Eitner, Der Gerber. Bd. XXIV. No. 563–567, 1898.
33. The Rationale of Drenching, by J. T. Wood. Leather Trades Review, Nov. 15, 1898.
34. Cantor Lectures on Leather Manufacture, by Prof. H. R. Procter, F.I.C., Society of Arts. Lecture II., April 24, 1899.
35. Further notes on the Action of the Dung Bate, by Joseph T. Wood. Jour. Soc. Chem. Ind. XVIII. 1899. p. 990.
36. Das Beizen der Blössen in Handbuch der Chromgerbung. Jos. Jettmar. Schulze and Co., Leipzig, 1900.
37. J. Borgmann. Die Feinleder Fabrikation, Berlin, 1901. Das Beizverfahren, p. 65.
38. Das Beizen der Blössen in Praxis und Theorie der Leder Erzeugung, von Josef Jettmar, Julius Springer, 1901.
39. Micro-organisms and Antiseptics in Tanning. F. Jean, Rev. Chim. Ind., 1901.
40. The Principles of Leather Manufacture, by Professor H. R. Procter. E. and F. N. Spon, London, 1903. Chap. XIII.
41. La Tannerie, par Louis Meunier and Clément Vaney, Gauthier-Villars. Paris, 1903. p. 125, etc.
42. Bakteriologische Vorgänge in der Lederindustrie. 1904, H. Becker. Zeitschrift f. Offentliche Chemie, Heft. XXIII. Jahrgang. X.
43. Untersuchungen über die Bakterien im Verdauungskanal des Rindes. P. Ankersmit. Centralbl. f. Bacter. I, O, Bd. 39, 1905. p. 359.
44. Über anaerobe Bakterien im Rinderdarm, Neubauer. Archiv f. wissensch. u. prakt. Tierheilk. Bd. 31, p. 153, 1905.
45. Studien über fäulnisserregende anaerobe Bakterien des normalen menschlichen Darmes, und ihre Bedeutung. F. Passini. Zeitsch. f. Hygiene. Bd. 49, p. 135. 1905.
46. A new process of puering or bating hides and skins. Allen Roger. J.S.C.I., p. 103, 1906.
47. Recent advances in the Bacteriology of Putrefaction, by J. T. Wood. J.S.C.I. XXV. 1906, p. 119. (Contains a bibliography of 21 items.)
48. Über den Bakteriengehalt menschlicher und thierischer Fäces. M. Lissauer. (Archiv f. Hyg. Bd. 58, p. 136.) See also Koch’s Jahresbericht, 1906, p. 195.
49. Character of the Bacterial Flora of Carnivorous and Herbivorous animals. A. Herter. (Science N.S. Vol. 24, p. 859. 1906.)
50. Leather Trades Chemistry. S. R. Trotman, Griffin, 1908. p. 79.
51. Le Rôle des Microbes dans la putrefaction des peaux en poils et en tripe et dans les confits, par Dr. Gr. Abt. Bull. Mens. du Synd. Gen. de cuirs et peaux de France. Nov.-Dec., 1908. p. 416.
52. Bakterien in der Lederindustrie. H. Becker, Collegium, 1909. p. 169.
53. A new Chromogenic Organism. S. R. Trotman, J.S.C.I., 1909. p. 1238.
54. Problems of the Leather Industry. Prof. H. R. Procter, J.S.C.I., 1910. p. 329.
55. Les Anaerobies, par M. Jungano et A. Distaso. Paris, Masson and Cie., 1910.
56. The Bacteriology of the Leather Industry. J. T. Wood, J.S.C.I., June 15th, 1910. p. 666.
57. The Seymour-Jones Anthrax Sterilization Method, by Alfred Seymour-Jones. Bradbury, Agnew and Co., Ltd., December, 1910.
58. Hauser, G. Ueber Fäulnissbacterien und deren Beziehungen zur Septicämie. Ein Beitrag zur Morphologie der Spaltpilze, 1885.
59. Woodhead, G. S. Bacteria and their products. (W. Scott, 1891.)
60. Carbone, Tito. Ueber die von Proteus Vulgaris erzeugten Gifte (Cent. Blat. f. Bakt., Dec. 4, 1890).
61. Macé, Traité pratique de Bacteriologie. Paris. Baillière.
62. Bienstock, Recherches sur la putréfaction. (Ann. de l’Inst. Pasteur, 13, 854.)
63. Klein. Ein Beitrag zur Kentniss der Leichenverwesung. (Cent. Blatt. l. 25, 278.)
64. Beijerinck, W. Schwefelwasserstoffbildung in den Stadt. gräben und Aufstellung der Gattung Aerobacter. (Central Bl. f. Bakt. II., Bd. 6, p. 193.)
65. Stich, C. Ueber die Bildung gasförmiger Phosphorverbindungen bei der Fäulniss. (Mitth. a. d. analyt. Laborat, der Krankenhausapotheke Leipzig, p. 22.)
66. Vitali, D. Bildung von Alcohol bei der Fäulniss von Proteinsubstanzen, die von Kohlehydraten befreit sind. (Bul. chimico farmaceutico, vol. 38, 1889, p. 729.)
67. Behrens, J. Fadenziehendes Brot. (Wochenbl. d. landw. Vereins im Grossherzogthum Baden, p. 569.)
68. Bienstock. Untersuchungen über die Aetiologie der Eiweissfäulniss, Milchfäulniss, Verhinderung der Fäulniss durch Milch, Darmfäulniss. (Archiv f. Hygiene, Bd. 39, 390.)
69. Taylor, A. E. Ueber Eiweissspaltung durch Bacterien, Zeitschr. f. Phys. Chemie, Bd. 36, p. 487.
70. Tissier, H., et Martelly. Recherches sur la putrefaction de la viande de boucherie. Ann. de l’Inst. Pasteur, t. 18, p. 865.
71. Wohltmann, F., Fischer, H., and Schneider, Ph. Bodenbakteriologische und Bodenchemische Studien aus dem Popplesdorfer Versuchsfelde. (Jour. f. Landwirthschaft, 1904, 97.)
72. Osterwalder, A. Ueber eine bisher unbekannte Art der Kernobstfäule, verursacht durch Fusarium putrefaciens nov. spec. (Centralbl. f. Bakt. 2 Ab. Bd. 13, p. 207.)
73. Schreiber, Karl. Fettzersetzung durch Microorganismen. (Archiv f. Hygiene, Bd. XLI. 328, Cent. Bl. IX. 849.)
74. Rahn, Otto. Die Zersetzung der Fette. (Cent. Bl. f. Bakt. Zweite Abth. XV. 53.)
75. Omelianski, W. Sur la fermentation forménique de la cellulose. (Arch. de sciences biol. St. Petersburg, t. 11, 251. Centr. Bl. f. Bakt. II., Bd. 8, 193.)
76. Dallinger and Drysdale. Monthly Microscopical Journal, vol. X., 53, 245; XI., 7, 69; XII., 261; XIII., 185; XIV., 106.
77. Lermer, K. Ueber die Produkte der Fäulniss der Gerste, Zeitschr. f. d. ges. Brauwesen, p. 165.
78. Salkowski and Neuberg. Die Verwandlung von d. Glukuronsaure in 1-Xylose Zeitschr. f. physiol. Chemie Bd. 36, p. 261.
79. Études chimiques sur la Végétation. J. Raulin, Paris, 1870.
80. Die Spaltpilze. W. Zopf. Breslau, 1885.
81. A Text Book of Tanning. H. R. Procter. Spon, 1885.
82. Micro-organisms and Disease. E. Klein. Macmillan, 1886.
83. Manual of Bacteriology. E. M. Crookshank. Lewis, 1887.
84. Manures, Natural and Artificial. W. Iveson Macadam. J.S.C.I., 1888, p. 79.
85. Methods and Formulæ used in the Preparation of Animal and Vegetable Tissues. P. W. Squire. Churchill, 1892.
86. Micro-organisms and Fermentation. A. Jörgensen. Macmillan (London), 1900.
87. Micro-organisms in Water. Percy Frankland and Mrs. Frankland. Longmans, 1894.
88. Disinfection and Disinfectants. S. Rideal. Griffin, 1895.
89. Guide to the British Mycetozoa, Dept. of Botany, British Museum. Arthur Lister, F.L.S., 1895.
90. Elementary Practical Bacteriology. Kanthack and Drysdale. Macmillan, 1895.
91. Les Ferments Solubles. E. Bourquelot. Paris, 1896.
92. The Manufacture and Applications of Lactic Acid. A. Claflin. J.S.C.I., 1897, p. 516.
93. Bakteriologie. C. Günther. Leipzig, 1898.
94. Bates and New Bating Materials. Der Gerber 24 [563], [564] and [565]. J.S.C.I., 1898, p. 361.
95. Les Enzymes. J. Effront. Paris, 1899.
96. Fixerung, Färbung und Bau des Protoplasmas. A. Fischer. Jena, 1899.
97. Soluble Ferments and Fermentation. J. Reynolds Green. Cambridge University Press, 1899.
98. Ueber die Zusammensetzungund Wirkungsweise der Mistbeizen. Der Gerber, 1899, p. 31.
99. The Mycetozoa. Sir Edward Fry and Agnes Fry, London, “Knowledge” Office, 1899.
100. Le Proteus Vulgaris. L. Felz. Baillière, Paris, 1900.
101. Bating Process, Improvements in. Leather Trades Review, 34 [773], 21. Abstr. in Jour. Soc. Chem. Ind., 1901, p. 263.
102. Oppenheimer, C. Ferments and their Actions. Trs. C. A. Mitchell. Griffin and Co., 1901.
103. Harden. The Chemical Action of Bacillus Coli communis on Carbohydrates and allied Compounds. J. Chem. Soc., Trans. 79, 1901, 610.
104. Traité de Technique Microbiologique. Nicolle et Remlinger. Doin., Paris, 1902.
105. Carini. Sull’ Applicazione della Bilancia idrostatica per il controllo della concia delle Pelli. Milan, 1903.
106. Traité d’Electrochimie. Max Le Blanc. Paris, 1904.
107. The Role of Diffusion during Catalysis by Colloidal Metals and similar Substances. Henry J. S. Sand, Ph.D. M.Sc. Proc. Chem. Soc. 74, 356. 1904.
108. Chemistry of the Proteids. G. Mann. Macmillan, 1906.
109. Canning and Preserving Food Products, with Bacteriological Technique. E. W. Duckwall, M.S., Bacteriologist to the National Canners Laboratory, etc. New York, 1905.
110. Précis de Coprologie Clinique. R. Gaultier. Baillière, Paris, 1907.
111. Allgemeine Chemie der Kolloide. A. Müller. Leipzig, 1907.
112. Zur Erkenntniss der Kolloide. R. Zsigmondy. Jena, 1905.
113. The Nature of Enzyme Action. W. M. Bayliss. Longmans, 1908.
114. Ueber die Interfibrilläre Substanz der Lederhaut bei Säugethieren. G. H. B. Van Lier. Coll., 1909, 321.
115. Practical Causerie on Deliming. Alex. T. Hough. Tanner’s Year Book, 1909, p. 83.
116. Grundriss der Kolloid Chemie. M. Ostwald. Dresden, 1909.
117. Beizen. W. Appelius. Technische Briefe Monatshefte von “Häute und Leder,” No. 23. April 1909.
118. Kolloid Chemische Beihefte. Ed. Wo. Ostwald. Dresden, 1909–1911.
119. Verfahren zum Beizen von Häuten. Otto Röhm. Coll., 1909, p. 58.
120. A Brief Review of Bacteriological Research in Phytopathology, with Bibliography. M. C. Potter, Sc.D., M.A. Science Progress, Oct. 1910, p. 191.
121. La Chimie de la Matière Vivante. J. Duclaux. Felix Alcan, Paris, 1910.
122. Sensibilité de la peau verte, et de la peau après l’échauffe, les pelains, et les confits, à l’égard de la chaux, du sel, et de l’acide acétique. Georges Abt et Edmund Stiasny. Coll., 1910, p. 189.
123. Bennett, H. G. The Analytical Examination of Bating. Leather Trades Review, 1911, pp. 697, 972.
124. Der Bacillus bulgaricus des Yoghurt in der Gerberei. Leder-technische Rundschau, No. 25, 1911. Collegium, 1911, p. 459.
125. Physiological Chemistry. Olaf. Hammarsten. Trans. J. A. Mandel, Wiley, 1911.
126. Précis de Biochemie. E. Lambling. Paris, 1911.
127. Bacteriological and Enzyme Chemistry, an Introduction to. Gilbert J. Fowler. Edward Arnold, 1911.
128. Microbes et Toxines. E. Burnet. Bibliothèque de Philosophie Scientifique. Flammarion. Paris, 1911.
129. The Physical Chemistry of the Proteins. T. B. Robertson, Professor at the Berkeley University, California, 1911.
130. Contributions to the History of the Enzyme Bate. Collegium, 1911. Otto Röhm, H. Becker, G. Eberle, J. T. Wood, pp. 271, 276, 278, 281, 312, 324.
131. The Fungi of Excreta. Jas. Scott. The Leather World, Jan. 11, 1912, p. 21.
132. Microbiology, by various Contributors. Edited by Charles E. Marshall, East Lancing, Michigan, Professor of Bacteriology and Hygiene, Michigan Agricultural College. J. and A. Churchill.
133. Technical Mycology. Lafar. C. Griffin and Co., 1911.
134. Manual of Bacteriology. R. T. Hewlett (Churchill), 1912.
135. Taschenbuch für den bakteriologischen Praktikanten. Dr. Rudolf Abel. Würzburg. A. Stuber’s Verlag.
List of Authors.
The numbers refer to the Bibliography.
Professor Procter, in his paper, entitled “Problems of the Leather Industry,”185 says: “There is, however, no reason that all the necessary effects both of puering and bating should not ultimately be attained by purely chemical treatment without the risk and uncertainty which must always attach to bacterial and ferment action.”
While I agree with him in this, still it is well to remember that in the case of one of the very oldest of the fermentation industries, that of the production of alcohol, a comparatively simple body, the natural process has not yet been replaced by a chemical one, and I believe this applies also to the manufacture of vinegar.
Chemical methods of bating may be used for leathers like chrome and alum leather, but even here natural processes like drenching, in which acids are produced gradually during the working of the skins, give more beautiful results. With vegetable tanning materials, the advantage is still on the side of the natural processes when these are conducted in a proper manner.
Finally, the author regrets that the subject of the work is not an inspiring and lofty one, such as is astronomy for example.186 Limited in extent, imperfect in execution, and in parts only suggestive in character, this little book may perhaps serve as a foundation, on which students of the science of tanning may raise the superstructure of their own experience and eventually perfect the processes touched upon, so that the use of such filthy materials may be entirely avoided. In that case the author’s intention will be fulfilled.
FINIS
Absorption phenomena, 70
Abt, Dr. G., 51, 72
Acid “turning” of drench, 237
Acids, action of, 156
— fractionating, 270
— deliming with, 9
— in drench, 265
— to remove lime, cost of, 11
Adulteration in dog-dung, detecting, 144
Agglutinin, 144
Alum, a term for puer, 3
“Alumed” calf skins, 3
Amend’s bate, 193
Amido compounds, 156
Amines, 163, 206
— in dung, 154
Ammonia in puer liquors, 41
Ammonium phosphate, 45
Analyses, bird excreta, 53
— dog dung, 26, 150, 152
— drench, 249
— gas, 247
— grains, 7
— guano, 150
— hen dung, 150
— puer, 26
— ultimate, of dung, 153
Andreasch, F., 244
Anti-bodies, 143
Anti-calcium, 236
Arginase, 133
Artificial drench, 263
Ash of skin, 39
Authors, list of, 290
Bacillus butyricus, 112
— c, 172
— Coli commune, 94
— d, 106, 173, 174
— e, 107, 173
— erodiens, 97
— from pigeon dung bate, 172
Bacteria, action of, 160
— bate, examination of, 87
— bran, pure culture of, 269
— dung, 89, 98, 102, 161
— growth in various media, 105
— pigeon dung, 103
— putrefactive, 125
— “Sweating,” 105, 213
Bacterial diastase, 131
Bacterium furfuris, 241, 264, 269
— termo, 118
Barium salts from pure cultures of B. furfuris, 271
— — of volatile acids, 253
Bate, Amend’s, 193
— Becker’s patent, 227
— bird dung, 52
— Boehringer’s patent, 197
— concentration of salts in, 47
— Dermiforma, 189
— Eberle’s patent, 221
— Erodin, 179
— Esco, 192
— examination of bacteria in, 87
— glucose, 179
— Lederer’s, 196
— Nordlinger’s, 195
— Norris’s, 194
Bate, Nowak’s patent, 194
— Oake’s, 195
— Oropon, 140, 186, 219
— phosphates in, 43
— pigeon dung, 103
— Phosphorbutyralin, 195
— Popp and Becker’s patent, 197
— Puerine, 190
— Purgatol, 192
— reducing action of, 152
— Röhm’s patent, 219
— Simon’s, 195
— solid matter, influence of, 176
— soluble matter of, 176
— Sucanine, 191
— Tiffany’s, 179
— Wood’s American patent, 210
— — English patent, 205
— Zollikoffer’s, 178
Bayliss, Dr. W. M., 127
Béchamp, 280
Betulase, 132
Becker’s patent, 227
Bibliography, 282
Bile, 31
— action of, 159
— colouring matters, 30
— salts, effect on puer, 46
— — — — lipase, 142
Bird dung bate, 52
— excreta, composition of, 53
Boehringer’s patent, 194
Bone meal, 196
Books of reference, 89, 282
Bran bacteria, 240
— — pure cultures of, 269
— composition of, 257
— drench, 233
— — non-volatile bodies in, 254
— — products of fermentation, 247
— — sweet, 263
— — volatile bodies in, 249
— ferments, action on starch, 258
Brown, Adrian J. Prof., 274
Butyric acid, 11
— — fermentation, 111
Calcium in enzymes, 136
— phosphate, 45
Carlsberg vessel, 170, 214
Caroubinase, 132
Casease, 132
Castor-oil seed, 188, 232
Catalysts, 127
Cellulose fermentation, 110
Cerealin, 245, 260
Chemical composition of puer, 24
— deliming, 5
Chloroform, use of, in pancreatin solution, 151
Cholesterol, 30
Collagen, 56
Colloidal state, 57, 137
Concentration of salts in bate, 47
Condenser, Abbe’s, 87, 88
— dark ground, 87
Conductivity of puer liquors, 80, 81
Copenhagen conference, 180
Cultivation of bating bacteria, 199
Culture medium, 105, 207
Cytase, 132
Deliming, chemical, 5
Density of skin, 60
Dermiforma, 189
Dextrin, 259, 261
Diastase, 128, 131, 132
— Krawkov’s method of preparing, 135
Diastatic enzymes, 235
Digestive ferments, 148
— juices, 230
Dog dung, analyses of, 26, 150, 152
— bacteria in, 89, 98
— detecting adulteration of, 144
— fresh, bacteria in, 102
— influence of food on, 28, 95, 153
— method of analysis, 25
Dog dung, separating enzymes from, 134
— trypsin in, 134
Doerr and Reinhardt’s laboratory, 32
Drench, acid “turning” of, 237
— acids in, 156
— action of, 233, 242, 245
— artificial, 263
— bacteria in, 239
— damage, 236
— fermentations of, 238
— gases from, 242, 248
— glucoses in, 261
— “pinholey,” 238
— putrid “turning” of, 237
— sweet bran, 235
Dressing hides, 19
Drop culture, 87
Dung analyses, 26, 150, 152, 153
— mineral matter of, 153
Eberle, G., 13
Eberle’s patent, 221
Effront, Dr., 11
Eitner, 242, 244
Electrometric apparatus, 76
— method, 33, 49
Emulsin, 132
Enzymatic bates, 197
Enzymes, action of, 145
— — of serum on, 144
— — on skin, 140
— calcium in, 136
— defined, 127
— diastatic, 235
— from bacteria, 131
— hydrolysing, 132
— in dog’s dung, 137
— lipolytic, 145
— manganese in, 136
— preparation of, 134, 164
— proteolytic, 145
Erepsin, 133, 142
Erodin, 179
Erodin, stock liquor, 182
Esco, 192
Esters, synthesis of, 127
Ethyl butyrate, 165
Ethylamine butyrate, 42
— lactate, 42
— propionate, 42
Fahrion, Dr., 52
Falling, 67
— apparatus for measuring degree of, 82
Fat in puer, 25, 30
— in skins, 16
Feeding of dogs, 28, 95
Fermentation in the leather industry, 147
— nitrogen from, 279
Ferments, digestive, 148
— gall, 224
— hydrolysing, 132
— imino-lytic, 133
— oxidizing, 133
— oxylytic, 133
Flagellæ, staining of, 94
“Flaked” grain, 149
Formic Acid, 9
Fractionating acids, 250
Frankland and Frew, 249
Gall ferments, 224
Gamgee, 281
Gas analysis, 247
Gases from drench, 242, 248
— from pure cultures, 275
— intestinal, 281
Gayon, 280
Gelatin, 56
— density of, 63
— glucose, 267
— molecular weight of, 56
— peptonized, 168
Glucose bate, 179
— gelatin, 267
Glucoses in drench, 261
Glycerin, 141
Goat skins, 183
— — bating of, 231
Guano, analysis of, 150
Golding, J., 175
“Grains,” analysis of, 7
Harness backs, 17
Hauff, J., 236
Hen dung, analysis of, 150
Hexoses, 125
Hide-powder, density of, 66
Hides, bating of, 17
Houston, Dr., 93, 94, 97
Hydrogen from drench, 239
Hydrogen-ion concentration, 74, 79
Hydrochloric acid, use in deliming, 11
Hydrolysis, 128
— velocity of, 130
Hydrolysing enzymes, 132
Immendorf, 280
Imino-lactic ferments, 133
Indol, 95, 106
Infusoria, 123
Intestinal gases, 281
— juice, 225, 228
Inulase, 132
Jean, F., 41
Jones, Arnold Seymour, 75, 133
Kaolin, 176
Kathreiner, F., 46, 236
— bate trials by, 183, 184
Koch’s Jahresbericht, 125
Körner, Dr. Th., 67, 167
Kossel, Dr. A., 170
— and Dakin, 133
Kral, Prof., xv
Kühl, Dr. H., 236
Laccase, 133, 135
— manganese in, 135
Lactase, 132
Lactic acid, 10
— — from B. furfuris, 272
Lamb, M. C., 9
Law’s volumenometer, 61
Lecithin, 141
Lederer’s bate, 196
Lime in sheep grains, 23, 37
— in puer liquors, 36, 40, 44
— soaps, 16
Limed skin, 5, 7
Lion’s dung, 29
Lipase, 132, 141
— effect of bile salts on, 142
Lipolytic enzymes, 145
Liquor pancreaticus, 189
Malase, 133
Maltase, 132
Manganese in enzymes, 136
Mastering, a term for puer, 3
Mercaptans, 46
Measuring truck, 8
Meggitt, Loxley, 171
Metchnikoff, 93
Mineral matter of dung, 153
Minot, Prof., 81
Mixed cultures, 173
Molecular weight of gelatin, 56
Monads, 123
Morocco leather, 3
Moulds, 114, 126
Müller, 72
Muscle, contraction of, 73
Myrosin, 132
Myxomycetes, 116
Nencki, 107
Nitrogen from fermentation, 279
— in bran, 234
— in puer liquors, 48
Nitrites, use of, in bating, 193
Non-electrolytes, influence of, 72
Nördlinger’s bate, 194
Nowak’s patent, 194
Nuclein, 114
Oake’s bate, 195
Oenoxydase, 133
Olease, 133
Organic acids, use in deliming, 13
— — in puer, 155
Oropon, 140, 186, 219
Osmosis, 58
Osmotic pressure, 59, 167
O’Sullivan, J., 244
— and Thompson, 128
Oxalic acid fermentation, 111
“Oxidizing” skins, 193
Oxydin, 133
Oxylytic ferments, 133
Paddle, 14
Pancreas, 220
Pancreatin, action on skin, 138, 151
Papain, 132
Parker, Dr. Gordon, xv, 11
Pectase, 132
Pentoses, 125
Pepsin, 132
— action on skin, 138
Peptolytic ferments, 119
Peptonized gelatin, 168
Phosphates in the bate, 43
Phosphorbutyralin, 195
Pigeon dung, 103
“Pinholey” drench, 238
Plasmase, 132
Plate cultures, 103
Popp and Becker’s patent, 197
Precipitin, 144
Pribram, Dr., 73
Procter, Prof. H. R., 10, 55, 205, 210
Proteolytic enzymes, 145
— ferments, 119
Proteus bacteria, 118
Puer, amido compounds in, 156
— chemical composition of, 24
— fat in, 25, 30
— organic acids in, 155
— quantity used, 15
— reactions of, 31
— wheel, 13
Puerine, 190
Puering, solution of skin substance in, 47
Pure cultures of bran bacteria, 269
Purgatol, 192
Purin bases, 30
Putrefaction, 116
— See Nos. 58 to 78 of Bibliography, 282
— mechanism of, 107
— of meat, 120
Putrefactive bacteria, 107
Putrid “turning” of drench, 237
Reaction of puer, 31
Rennet, 128, 132
Rhamnase, 132
Rogers, Dr. Allen, 195
Röhm’s patent, 219
Ruge, 281
Sabatier and Senderens, 127
Salt solution, physiological, 87
Salts solution, 267
Sand, Dr., xv, 33, 81
Schorlemmer, K., 186
Scud, 51
Scudding, 16
— machine, 17
Serum, action of, on enzymes, 144
Shearling pelts, 183
Shrewsbury, H. S., 166, 281
Simon’s bate, 195
Skatol, 46
Skin substance, ash of, 39
Skin substance, density of, 60
— — fat in, 16
— — solution of, 47, 51
Soerensen, Prof., 43, 80
Solid matter in bate, 73
— — influence of, 175
Soluble matter of bate, 176
Soyka’s dilution method, 103
Spirillum volutans, 112
— delsulfuricans, 124
Spongy leather, 238
Staining and mounting, books on, 89
— of flagellæ, 94
Starch, action of bran ferments on, 259
Steapsin, 132
Stiasny, Prof. E., 282
Sucanine, 191
Sucrase, 128, 132
Sulphuric acid, use in deliming, 11
Sulphuretted hydrogen, 46
“Sweating” bacteria, 105, 213
Sweet bran drench, 235
Swelling, 67
— alkaline, 72
Symbiosis, 269
Synthesis of esters, 127
Syntonin, 32
Thio-acids, 196
Tiffany’s bate, 179
Toxins, 143
Trehalase, 132
Trent bridge laboratory, 107, 252
Trimethyl-amine, 107, 252
— butyrate, 42
Trypsin, 132, 133
Trypsin, in dog dung, 134
Turney, Sir John, scudding machine, 17
“Turning” of drench, 287
Tyrosinase, 133
Urea, 29
Urease, 132
Uric Acid, 29
Velocity of hydrolysis, 130
Volumenometer, 61
— air, 67
— Law’s, 61
Wladika, J., 256
Willcox, Dr. W. H., 243, 245
Wood’s American patent, 210
— English patent, 205
Wool bacteria, 174
Xylose, 125
Yeast water, 259
Yoghurt, 236
Zinc, effect on growth of moulds, 113
Zollikoffer’s bate, 178
Zoogloæ, 242
Zymase, 133
LONDON: PRINTED BY WILLIAM CLOWES AND SONS, LIMITED
GREAT WINDMILL STREET, W., AND DUKE STREET, STAMFORD STREET, S.E.
1 Report of First Conference of Leather Trade Chemists, 1897, p. 146.
2 Jour. Chem. Soc., Mar. 1886.
3 Jour. Soc. Chem. Ind., 1894. p. 218.
4 Died, April 1905.
5 There is a copy in the British Museum and also in the Patent Office Library.
6 Some further analyses of grains at Trent Bridge gave the following results:—
Water | CaO | CaO on dry skin | |
| Grain from splitting machines | 76·3 | 1·14 | 4·8 |
| The same after washing 24 hrs. | 86·0 | 0·22 | 1·57 |
| The same after puering | — | — | 0·80 |
| Another grain | 79·7 | 1·06 | 5·20 |
| The same after deliming as completely as possible with HCl | — | — | 0·45 |
| Skin in natural condition | 64·0 | — | 0·125 |
M. C. Lamb has found in sheep grains split for skivers the following amounts:—
Ash | CaO | CaO in Ash | |
| Grain from splitting machine | 9·3 | 3·2 | 34·8 |
| The same after washing | 4·1 | 1·5 | 35·1 |
| The same after puering | 3·1 | 0·9 | 29·1 |
| The same after bran drench | 1·7 | 0·55 | 32·4 |
The figures are all percentages calculated on the dry weight of the skin.
Munro Payne (Tanners’ Year Book, 1905, p. 75) gives the following amounts of lime as Ca(OH)2 in limed hides, calculated on dry weight at 212° F.:—
| Direct from limes | max. | 3·859 per cent. | |
| " " | min. | 2·836 | " |
| Limed for buff | 4·621 | " | |
| Limed for tanning | 3·7659 | " | |
| Ditto after bating | 0·689 | " | |
| Calf limed | 2·601 | " | |
| Calf bated | 0·1215 | " | |
| Goat limed | 5·613 | " | |
| Goat bated | 1·268 | " | |
7 From the molecular weights it will be seen that 74 grm. of calcium hydrate, equivalent to 56 grm. of lime (CaO), require 73 grm. of HCl gas for neutralization. This quantity of gas is contained in 265 grm. of commercial hydrochloric acid of 18° Bé., or about 230 c.c.—that is, for 100 grm. CaO, 410 c.c. of 18° Bé. acid are required; hence for the 100 kilos wet grain containing 400 grm. CaO, 1640 c.c. of acid are required for complete neutralization. It is impracticable to use this amount of acid, for the reasons stated above.
8 Tanners’ Year Book, 1911, p. 59.
9 See Chapter III.
10 See also L’Acide Butyrique dans la Tannerie, Urbain J. Thuau, Le Cuir, Aug. 1910. Also Collegium, 1910, pp. 347, 363.
11 Le déchaulage des Peaux en tripe, Ettore Guisiana (Turin), Coll. 1910, p. 14.
12 Verfahren zum Beizen von Haüten, Ledertechnische Rundschau, No. 24, 1910. Coll. 1910, p. 372.
13 When the puer is dirty it should be diluted with water and strained through a bag, or the diluted puer may be put into a tub and the heavy grit and dirt allowed to fall to the bottom of the vessel. Puer from hunting kennels is usually clean enough to use without this process.
14 The quantities of puer used vary considerably in different works. At Trent Bridge, 10 to 15 lb. of wet puer from hunting kennels are used per 100 lb. of wet pelt. In a German works, 15 kilos of fairly dry dog dung are used per 100 kilos of pelt (Blösse). In an American works, two buckets (one bushel) wet dog dung were used to puer 10 dozens of 8 lb. calf skins. This is equal to about 62 lb. of puer to 100 lb. of skin, which seems an excessive quantity. In an Italian works, two kilos dry dung were used to 20 skins of sheep, equivalent to about four kilos per 100. In an English tannery, 100 hides for harness leather required four bushels by measure of pigeon dung, costing 1s. 6d. per bushel (one bushel equals eight gallons).
15 A German firm, who supply dry dog and pigeon dung for bating purposes, recommend that the material should be spread in a warm room at 30° C., and moistened up to 80 per cent. of its weight with an infusion of oat straw, and continuously turned over until completely softened. Other materials may be used for the fermenting fluid in place of the oat straw infusion, such as broth made from waste fleshings, whey, or even a simple inorganic solution, such as is used for cultivating bacteria, consisting of—
2 parts potassium phosphate.
1 part magnesium sulphate.
1 part calcium nitrate.
1·5 part calcium chloride.
100 parts water.
This procedure is neither more nor less than an application of the principles of bacterial cultivation and propagation, shown to be practically useful in the bating process by Professor H. Becker, of Frankfort, and by the Author in various publications (see Chapter VII.).
16 There is considerable variation in the length of time required to bring down the skins. Lamb states that half an hour is frequently sufficient, but at Trent Bridge 2–3 hours are generally required. The causes of the variation are not altogether clear, but depend partly on the previous history of the goods (fellmongering, liming, etc.), and partly on the water used.
17 The original Turney scudding machine, was invented in 1871, and patented in that year. The specifications are numbered 1351, and 3310, 1871. It was also patented in America. Specification No. 131,480, dated September 17, 1872. Improvements in apparatus for cleaning and scouring hides and skins.
18 Limed sheep grains, or Pelts, which have been limed for splitting, contain from 3 per cent. to 6 per cent. CaO on the dry skin (see footnote, p. 7), and about 80 per cent. of water. In a typical case of a fully limed grain, the lime was found to be distributed as follows:—
| Free uncombined lime (CaO) | 1·7 | per cent. on the dry skin |
| CaO combined with skin | 2·5 | " |
| CaO as carbonate, or other salts | 1·4 | " |
── | ||
5·6 |
Thus, 75 per cent. of the total lime in the skin was in the caustic state. If the skin be cut during the progress of the bating operation, and a solution of phenolphthalein applied to the cut surface, the course of bating may be followed, and it will be found that the caustic lime disappears from the skin in a very short period of time, accompanied by the sudden collapse of the fibres. We shall treat of this more fully in Chapter III. After puering, the lime (CaO) contained in the skin, amounts to 0·5 to 0·9 per cent., calculated on the dry skin. This lime is in a perfectly neutral state, and is more or less in combination with the fibre of the skin. The exact state in which the lime exists in the skin after puering is not known, and would form a very interesting subject for research.
19 Schmidt, and Strassburger, Die Fæces des Menchen (Hirschwald, Berlin), 1901.
20 Lambling, Précis de Biochimie, 1911, p. 221.
21 Another sample contained 136 grm. dry matter.
23 Abderhalden, Physiological Chemistry.
24 Cf. Mann. Chemistry of the Proteids, 1906, p. 432.
25 The cholesterol was obtained by extracting the dry puer with ether, saponifying with alcoholic potash. Shaking out the unsaponifiables with ether, evaporating to dryness. The residue was taken up with absolute alcohol, decolorized with animal charcoal. On cooling, characteristic crystals of cholesterol separated. Hoppe-Seyler finds from 1 per cent. to 4 per cent. cholesterol in fæces. See also Gaultier, Coprologie Clinique, p. 160.
26 For qualitative tests for the colouring matters, see Gaultier, Précis de Coprologie Clinique, p. 159.
28 Probably syntonin, or bodies of a similar nature (see Allen. Comm. Org. Anal. iv. p. 4). The supposition is supported by the fact that solutions of puer have a slight laevorotatory action on polarized light.
See also note on Van Liers paper, Coll. 376, 1909, p. 823.
29 See paper entitled, “The Employment of the Electrometric Method for the Estimation of the Acidity of Tan Liquors,” by H. J. S. Sand, Ph.D. D.Sc., and D. J. Law, B.Sc. F.I.C., J.S.C.I. 1911, p. 3; also Part II., by Wood, Sand and Law, J.S.C.I. 1911, p. 872.
30 Eitner, Der Gerber, 566, p. 77, states that little or no neutralization of lime by acids or acid salts takes place in the bate.
31 Ber. Deutsch. Botan. Ges. 1900, xviii. 32, J.S.C.I. 1900, 386.
32 Centr. Bakt. Parasitenk, ii. Abt. viii., 393, etc., J.S.C.I. 1902, 1151.
33 Fitz (Berichte, 1876–1884, see Herfeldt, J.S.C.I. May 31, 1895), has shown that lactates in dung are decomposed by bacteria, the chief product being propionic acid.
34 Rettger, Amer. Jour. of Physiol., viii. p. 284; Koch’s Jahresbericht, 1903, p. 112.
35 Deliming with acids when carefully done takes out more lime than the puer, without in any way injuring the skin, but when tanned with sumac the bellies and axilla are harsh, and of a browner colour than in a puered skin. Analysis showed no more lime in the brown portions than in the centre of the skin, where the colour was quite bright and satisfactory.
36 Vide Procter, L.I.L.B., 1908, p. 64.
37 In a blank experiment to determine this, 0·0476 grm. N per litre was found to be brought into solution from the puer.
38 Stiasny, On Old Limes. Collegium 1910, p. 181.
39 Stiasny. Collegium 1910, p. 184.
40 Ueber die Zusammensetzung des beim Beizen von Lammfellen mit Hundekot abfällenden “festen Schmutzes”: ein Beitrag zur Beizenfrage.—Coll., 1911, p. 445.
41 Urea is present to the extent of 5 per cent. in samples of Peruvian guano.
42 Manures, Natural and Artificial, W. Ivison Macadam. Jour. Soc. Chem. Ind. 1888, p. 79.
43 Principles of Leather Manufacture, p. 179.
44 Stiasny, Kritische und Experimentelle Beiträge zur Aufklärung der Gerbvorgänge. Collegium 1908, p. 117.
Wood, Compounds of Gelatin and Tannin. Collegium 1908, p. 494.
45 Allen, Comm. Org. Analysis, iv. p. 459.
46 Procter, Colloidal Chemistry. Brit. Assoc. Rep. 1908.
47 M. Planck, Zeit. f. Phys. Chem. xlii. p. 584 (1903).
48 Sull’ appliccazione della bilancia idrostatica per il controllo della concia delle pelli, Milan, 1903.
49 See Kohlrausch, An Introduction to Physical Measurements, 1894, p. 43. See also, Weighing Hides in Water, C. E. Parker and G. H. Russell, Tanners’ Year-Book 1905, p. 45.
50 Wiedemann’s Annal d. Physik, xxxv. p. 352 (1888).
51 Th. Körner, 10 Jahres. Bericht, d. Deutsch. Gerberschule zu Freiberg, 1899, p. 32.
52 Procter, Ueber die Einwirkung Verdünnter Sauren und Salzlösungen auf Gelatine, Kolloid Chem. Beihefte, 1911, ii. p. 243.
53 Duvernoy, E. Wiedemann, and Lüdeking, Wied. Ann. xxv. 1885, p. 145.
54 Rodewald. Thermodynamik der Quellung, Zeit. f. Phys. Chem. xxiv. 1897, p. 193.
55 Zur Lehre von d. Quellung, Wied. Ann. liii. 1894, p. 564.
56 Nernst and Schönfliess. Einleitung in die mathematische Behandlung der Naturwissenschaft, München 2 Aufl. 67.
57 See also Colloidal Chemistry, H. R. Procter, M.Sc., Brit. Assoc. Reports, Dublin, 1908.
58 Wilhemy. Pogg. Ann. cxix. pp. 121, 122.
59 Colloidal Chemistry, p. 19.
60 Allgemeine Chem. d. Kolloide, p. 13.
61 Přibram, Die Bedeutung der Quellung und Entquellung für physiologische und pathologische Erscheinungen. Kolloid Chem. Beihefte, ii. p. 1.
62 Perrin, Kolloid Chem. Beihefte, i. p. 221.
63 A particle of gamboge in a colloidal solution has a mass 109 times that of a molecule of hydrogen.
64 Acids in Tan Liquors, Journ. Soc. of Chem. Ind., 1910, p. 1354.
65 Journ. Soc. Chem. Ind., 1911, p. 3.
66 Ibid., 1911, p. 872.
67 An improved form of hydrogen electrode devised by Dr. H. Sand may now be obtained from Universitäts-mecanikes Fritz Köhler, Leipzig. It is shown at I in Fig. 10.
68 It is necessary to use pure hydrogen. This may now be obtained commercially from the Knowles’ Oxygen Co., Wolverhampton.
69 The formula for calculating p+H for the N/1 auxiliary electrode used is as follows:—
Since π = (0·283 + 0·0575 log 1C volt)
log 1C = p+H = π – 0·2830·0575
π = potential difference found
C = concentration of hydrogen-ions.
70 Vide Sœrensen, Sur la mesure et l’importance de la concentration des ions hydrogène dans les réactions enzymatiques. Comp. rend. Lab. de Carlsberg, 8me. vol. 1ere livraison, Copenhagen, 1909.
71 Minot, Address to Amer. Assoc. for Advancement of Science, Minneapolis, Dec. 29, 1910. Nature, 1911, p. 96.
72 The apparatus was constructed by Mr. W. Linney, of the University College, Nottingham.
Since this apparatus was constructed Professor H. R. Procter has informed me that an appliance devised by Mr. Anderson, of the Leeds University, for measuring the thickness of leather under varying pressures, has been in use in the leather industries department for some months for the purpose of measuring the proportion of the permanent to elastic compression.
73 A good microscope, with 112 oil-immersion objective and Abbe condenser, is necessary for bacteriological work. The new dark-ground condenser made by James Swift and Son, London, is extremely useful for examining living bacteria under high magnifications.
74 The most recent method is to preserve the slides without cover-glasses: a drop of cedar oil is placed directly on the preparation, and, after examination, carefully washed off with xylol. Care should be taken not to touch the preparation with the fingers.
75 Masson et Cie, Publishers.
76 Klein, E., Micro-organisms and Disease (Macmillan), p. 22. See also, Kanthack and Drysdale, Practical Bacteriology (Macmillan 1896), p. 86.
77 The subject is not so far removed from direct human interest as one might suppose. For many years Metchnikoff, in Paris, has been studying the bacterial flora of the human intestines. His theory is, that old age is caused by the poisonous products of these intestinal bacteria, and he proposes to counteract the effects of these poisonous organisms, by introducing into the system large quantities of lactic acid bacteria, either in the form of tabloids, or in the form of sour milk cultures.
78 For the demonstration of flagellæ, the material is taken from a culture on solid media—a young agar culture is best. Make a dilution by Soyka’s method (p. 103), dry and fix on a cover-glass as described, p. 88, then proceed as follows:—
1. Mordant with a mixture of 10 c.c. 20 per cent. tannin solution, 5 c.c. cold saturated ferrous sulphate, 1 c.c. fuchsin or wool-black solution. This solution is put on the preparation, which is then heated for one minute until steam is given off.
2. Wash under the tap, then in alcohol.
3. Stain with anilin-water-fuchsin solution, prepared by dissolving the dye in anilin-water and adding 1 per cent. of a 1 per cent. solution of caustic soda until the liquid begins to go cloudy.
4. Wash off the stain with distilled water.
79 “Beitrag zur Bakterienflora des Darmes,” Archiv f. Hygiene, Bd. 26, p. 293.
80 Zeit. f. Offt. Chem., Heft. xxiii. Jahr. X. p. 447.
81 Dr. A. C. Houston has been kind enough to make an examination of B. erodiens, and to ascertain its action on various sugars. It produces fluorescence in neutral-red broth cultures, acid and gas in lactose cultures, indol, and acid, and clot in milk cultures. It ferments dulcite with production of acid, but not cane sugar, adonite, inulin, inosite, salicin, or raffinose.
82 The modification of Soyka’s method of making attenuations of bacteria is that of Günther (Bakteriologie, 1898, p. 204), and is carried out as follows:—On the inner surface of the cover of a sterile Petri dish (which is to be used for making the plate culture), place four drops of sterile broth, or sterile water; inoculate the first drop, by means of a platinum needle, with the material to be examined; heat the needle in the flame, dip it into the first drop, and, with the liquid adhering, inoculate the second drop. Proceed in this way, each time heating the needle, to the fourth drop. From this fourth drop a tube of nutrient gelatin is inoculated, and poured on to the plate. The cover is then put on, and the plate put into the incubator. The drops on the cover do not in any way contaminate the culture.
83 Villon found that Bacterium pilline developed 0·142 per cent. ammonia in ten days in lime liquors used for unhairing skin.
84 Cultures made from old limes have, however, not been found effective in bating.
85 Journ. f. Prakt. Chem., Bd. xvii., 1878; see also Stoklasa, Cent. Bl. f. Bakt. vi., p. 526.
86 Proc. Roy. Soc., lxvii., 1900.
87 Reynolds Green, Fermentation, p. 350.
88 Sensibilité de la peau verte, et de la peau après l’échauffe, les pelains, et les confits, à l’égard de la chaux du sel, et de l’acide acétique. Georges Abt et Edmund Stiasny, Collegium 1910, p. 1 9.
89 See Les Anaerobies, Jungano and Distaso (Masson et Cie Paris, 1910).
90 The Bacteriology of the Leather Industry, J.S.C.I., 1910, p. 666.
91 Reprinted from the Journal of the Society of Chemical Industry, February 15, 1906, No. 3, vol. xxv. The numbers in brackets refer to the Bibliography, Chapter XI.
92 J.S.C.I., 1894, 218.
93 J.S.C.I., 1893, 442.
94 “The Nature of Enzyme Action,” by Dr. W. M. Bayliss, FRS.
95 The important discoveries of MM. Paul Sabatier and Senderens on the catalytic action of finely divided metals, notably nickel and copper, have recently been extended to a study of the catalytic action of various metallic oxides. In the Comptes rendus, MM. Paul Sabatier and A. Mailhe give an account of a new synthetic method, based on the catalytic effect of titanium oxide, which would appear to possess many practical applications. They show that if a column of titanium dioxide is maintained at a temperature of 280°–300° C., and a mixture of the vapours of a primary alcohol and a fatty acid (other than formic acid) is led over it, the corresponding ester is formed. The same limit is here reached instantaneously as was found by Berthelot after prolonged contact. An excess of either constituent favours the limit of combination of the other. Following this method, the methyl, ethyl, propyl, butyl, isobutyl, and isoamyl esters of acetic, propionic, butyric, isobutyric, isovaleric and caproic acids have been prepared. Esters of benzyl alcohol have also been readily obtained by this method. The inverse action—the direct hydrolysis of esters by water—is also easily effected, and the use of titanium dioxide reduces any secondary reactions to a negligible amount. See Nature, March 9, 1911, p. 54. See also Dr. Sand’s paper—bibliography.
96 There are apparent exceptions and complications of this law which we shall not here enter into, except to say that they may be explained by the fact that the action of some enzymes is reversible (see p. 141, under lipase.)
97 “Zeit. f. Hygiene,” Bd. xlix. p. 135.
98 Les Enzymes, Dr. Jean Effront, Paris, 1899.
99 Zeit. f. Physiol. Chem. xli. f. 153 (1904). See also the articles by Mr. A. Seymour Jones, B.Sc., writing under the pseudonym of “Heof Joppa,” in the Leather Trades’ Review, July 19, 1911, p. 540; and Aug. 16, 1911, p. 625.
100 See, however, the recent paper by Eberle and Krall, Ueber den Nachweis des Trypsins im Hundekot, Collegium 1911, p. 201, in which the authors endeavour to show the presence of unchanged trypsin in dog dung. Their proof depends upon the action of an antipancreatic serum on infusions of dog dung based on the work of Achalme, Ann. de l’Inst. Pasteur, 1901, p. 737. See also the criticism on this paper by Dr. Otto Röhm and Dr. Max Goldman, Collegium, 1911, p. 265. Hammarsten (Physiol. Chemistry, 1911, p. 494) states that “among the secretions which undergo putrefaction in the intestine, the pancreatic juice, which putrefies most readily, takes place first.”
101 Duclaux. See Bibliography.
102 Koch’s Jahresbericht über Gärhungs Organismen, 1908, 635.
103 J.S.C.I., vol. xiii., 1894, p. 218.
104 The word pancreatin is used throughout in the sense of pancreas extract. As is well known, this contains several enzymes, trypsin, steapsin, maltase, and also a rennin.
105 Also known as steapsin, or pyolin (Allen, p. 357, vol. iv., Comm. Org. Analysis).
106 In the digestion of fat by dogs the glycerol produced is all absorbed before the ileum is reached, so that none exists in the excrement. (Levites, Chem. Soc. Abst. 1907, vol. iv. p. 891.)
107 Another instance of the reversibility of enzyme action is the secretion of a peptolytic enzyme, by B. pyocyaneus; i.e. it has the power of synthesizing proteins, as well as of decomposing them. (Zak. Chem. Soc. Abst. 1907, p. 996.)
108 Chem. Soc. Annual Reports IV., 1907, p. 252.
109 Hammarsten, p. 70.
110 Gulland, Ency. Britt. iv., p. 83.
111 Appelius, Technische Briefe, No. 23, April 1909.
112 Harden, see Bibliography.
113 References to bating:
Procter. Text-book of Tanning, 1885, 184.
Der Gerber, x. (1884), 197, Mistbeizen.
Der Gerber, xv. (1889), 267.
J. Anal, and Appl. Chem., 1893, vii. 87, 95.
J.S.C.I., xii. 530, Palmer and Sandford.
Salomon, W. J., Tech. Quarter, 1892, v. 81, etc.
J.S.C.I., xii. 774, ix. 27.
114 Harris and Gow, Jour. of Physiol., xiii. 469. G. Tammian, Zeitschr. f. Physiol. Chem., xv. 271. O. Loew., J. Prak. Chem., 37, 101. J.S.C.I., 1888, 224.
115 This is not invariably the case: other samples of puer examined have given off NH3 when similarly treated, but only in small quantity. A similar instance has been observed by Rideal and Orchard in the liquefaction and decomposition of gelatin by B. fluorescens liquefaciens (The Analyst, October 1897), where the quantity of ammonia produced was insignificant, amounting even after 16 days’ incubation to only 0·168 grm. of N per 100 c.c., corresponding to 0·204 grm. of ammonia.
116 Procter, Text Book of Tanning, p. 85.
117 According to the text books; but for later researches see Paal and Schilling, Chem. Zeit. 1895, 1487; also J.S.C.I., 1898, 589.
118 H. K. Procter, “Text Book of Tanning,” 1885, p. 18.
119 See Procter, “Text Book of Tanning,” pp. 8 and 21.
120 Since the above was written, Severin (Centr.-Bl. iii. [2], 628) has described seven other species of bacteria, isolated from horse dung; for the description and property of these the original memoir must be consulted. Popp and Becker, German Patent 86,335, 28, describe fourteen species which they isolated from dog and pigeon dung, of which three have considerable bating action on skins, and which form the subject of the patent. Some of these are probably identical with organisms in the above list.
121 Centr.-Bl. ii. [1], 97.
122 J.S.C.I., 1894, March 31, 218–221.
123 See Gamgee, Phys. Chem. 2.
124 Ann. de Micrograph, ii. [6], March 20, 1896.
125 I did not consider it necessary to purify the enzymes, the object being to examine the action of the bacterial products apart from the living organisms.
126 Reprinted from the Journal of the Society of Chemical Industry, November 30, 1899, No. 11, vol. xviii.
127 10 Jahresber. d. Deutsch. Gerberschule zu Freiberg, 1898–99: Beiträge zur Kentniss der wissenschaftlichen Grundlagen der Gerberei, p. 32.
128 J.S.C.I., 1898, p. 1011, see previous paper, p. 152.
129 Ber. 25, 1202; also Allen, “Comm. Org. Analysis,” iv., 466. F. Marpmann, Centralblatt f. Bakt. 2, 5, 67.
130 Rev. Gen. des Sciences, 1899, p. 380.
131 Jorgensen, the “Micro-organisms of Fermentation,” p. 20.
132 J.S.C.I., 1898, p. 1012.
133 Leather Industries, Sept. 1898.
134 Principles of Leather Manufacture, p. 160.
135 Text Book of Tanning, 1885, p. 185.
136 The Manufacture of Leather, Philadelphia, 1885.
137 At the third Conference of the I.A.L.T.C., held in Copenhagen Aug., 1899, F. Kathreiner (Worms) brought forward communications from Wood (Nottingham) and Dr. Becker (Frankfort-on-Main) on artificial bates, from which it is shown by experiments and trials on a large scale at the works of Messrs. Doerr and Reinhart that both these gentlemen had worked out similar processes quite independently. Wiss. Techn. Beilage des Ledermarkt, 1899–1900, p. 8. Bemerkungen über die Wirkung der Kothbeizen, J. T. Wood. Idem, p. 43.
Popp and Becker’s und Wood’s Ersatzmittel für Köthbeizen. Franz Kathreiner. Idem, p. 50.
Einiges über die Anwendung von Erodin einem Ersatzmittel für Hundekoth u. dergl. in der Leder industrie. Bericht von Dr. H. Becker, Frankfort-on-Main. Wiss. Techn. Beilage des Ledermarkt. Bd. i., p. 39.
Further particulars about the bate and its construction will be found in Chapter VIII.
138 One litre of erodin stock liquor (1 part erodin, 40 parts water) were used per kilo drained washed pelt, in the drum for 1 12 hours. In some cases less than 1 litre sufficed.
140 J.S.C.I., 1898, p. 1013.
143 A most useful source of information is Section XV. (Journal and Patent Literature) in the Journal of the Society of Chemical Industry, published fortnightly at Palace Chambers, Westminster. Abstracts of all patents relating to bates will be found under the above heading.
144 J.S.C.I. 1906, p. 103.
145 J.S.C.I. 1898, pp. 1010 to 1013, and 1899, pp. 990 to 993.
146 An abstract of Wood’s work. 1901, p. 148, section 4, p. 149.
147 1. See German Patent No. 200,519 or British Patent No. 537708 Roehm, Esslingen.
148 1. See German Patent of Addition No. 203,889 Roehm, Esslingen.
149 2. See Abderhalden, “Text-book of Physiological Chemistry,” 1906, page 555 and following.
150 1. See Abderhalden, “Text-book of Physiological Chemistry,” 1906, page 556.
151 Bran examined by Kjeldahl’s process contained 2·2 per cent. N, equivalent to about 13–14 per cent. proteids.
152 Principles, pp. 166–170.
153 Der Gerber, 1898, p. 204.
154 Claflin (92) gives the limits of acidity for lactic bacteria from 0·02 to 0·5 per cent. Reynolds Green (97, p. 346) gives the upper limit for B. acidi lactici as 8 per cent. lactic acid.
155 Der Gerber, 24, 570.
156 Jour. Soc. Chem. Ind., 1893, p. 426.
157 Der Gerber, 1895, p. 193, etc.
158 J.S.C.I., 1897, p. 513.
159 Ibid., 1890, p. 28.
160 Ibid., 1893, p. 422.
161 J.S.C.I., 1890, 27.
162 Zeits. für Biologie, xxiv. 105.
163 Jour. Chem. Soc., 1892, Trans. 254.
164 Der Gerber, xvi. (368), 4.
165 Method recommended to us by Dr. Frankland. Vide Jour. Chem. Soc. Trans. 59, p. 94. Appendix II. Determ. of Vol. Acids by dist. with HCl.
166 Julius Wladika. Vide Der Gerber, xvi. 28. Zur Kenntniss der Organ. Säuren in Fichtenbrühen.
167 J.S.C.I. 1890, 27.
168 7 grm. of yeast boiled in 100 c.c. H2O.
169 Prepared by precipitation with alcohol.
170 Prepared by heating starch and water to 50° C. for 18 hours and filtering.
171 Watts’ Dict., Old Ed. Cerealin discovered by Mèje Mouriés, Comptes rend. 37, 351; 38, 505; 43, 1122; 48, 431; 50, 467.
172 Vide der Gerber xiv. 257 Süsse Kleienbeize.
173 J.S.C.I. 1893, 422.
174 Potassium phosphate, 1 grm.; magnesium sulphate, 0·2 grm.; calcium chloride, 0·1; water, 1000 c.c. Vide Frankland and Frew, Trans. 1892, 255.
175 J.S.C.I. 1890, 28.
176 J.S.C.I. 1892, 422.
177 Vide Note on the Estimation of Butyric Acid, W. H. Willcox, J. Chem. Soc., Nov. 21, 1895.
178 Vide J. Chem. Soc. (Trans.), lix., 94, App. II.
180 2000 c.c. gave only about the same amount of acids as 1000 c.c. fermentations. For percentages of acids, see table, p. 281.
181 J.S.C.I. 1893, 423.
182 Schutzenberger, “Fermentation,” 1876, p. 227.
183 Woodhead, “Bacteria and their Products,” p. 125.
184 Die Stickstoffkonservirung im Stalldünger, Jour. f. Landwirthsh., xlii. p. 69.
185 J.S.C.I. 1910, p. 331.
186 Celestial Objects, Webb, 1881.
End of the Project Gutenberg EBook of The Puering, Bating & Drenching of
Skins, by Joseph Turney Wood
*** END OF THIS PROJECT GUTENBERG EBOOK PUERING, BATING, DRENCHING OF SKINS ***
***** This file should be named 58168-h.htm or 58168-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/8/1/6/58168/
Produced by Thiers Halliwell, Chris Curnow and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive)
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.